

Thin Mouse Shrew
Myosorex tenuis

2025 Red list status
Endangered
Regional Population Trend
Declining
Change compared
to 2016
No Change
Overview
Myosorex cf. tenuis – Thomas & Schwann, 1905
ANIMALIA – CHORDATA – MAMMALIA – EULIPOTYPHLA – SORICIDAE – Myosorex – tenuis
Common Names: Thin Mouse Shrew (English)
Synonyms: No Synonyms
Taxonomic Note:
Roberts (1951) recognised Myosorex tenui (Thomas & Schwann 1905) as a distinct species based on small cranial size. Although the type specimen is from the Zuurbron, Wakkerstroom District in the Mpumalanga Province, he also assigned M. tenuis to specimens from Woodbush and Entabeni Forests in the Soutpansberg Mountains, Limpopo Province. The species was not accepted by Meester et al. (1986) or Skinner and Chimimba (2005), but was tentatively accepted by Jenkins and Churchfield (2013). More recently, Willows-Munro (2008) and Taylor et al. (2013), using a combination of molecular and morphological characters, demonstrated considerable lineage diversification within the M. cafer complex. Isolated populations of M. cafer in the highlands of Zimbabwe and Mozambique; and north-eastern Limpopo have been previously assigned to M. cafer (Friedmann & Daly 2004). However, Taylor et al. (2013) described two new species from specimens previously assigned to these M. cafer populations: The Zimbabwe and Mozambique population are considered M. meesteri and the Limpopo lineage was tentatively assigned M. cf. tenius based on its small cranial size. However, it is unclear if the Mpumalanga specimen named M. tenuis by Roberts (1951) or the Mpumalanga records of M. cafer should be reassigned to M. cf. tenuis or remain M. cafer. Further vetting of museum records is needed to resolve the taxonomy. In order to finally resolve the suitability of M. tenuis as the correct name for the Limpopo lineage, further research is needed based on detailed comparisons of dental, morphometric, and molecular characters of the holotype of M. tenuis. Pending such analysis, Taylor et al. (2013) provisionally assigned the Limpopo populations to M. cf. tenuis. Whether the final name of the species changes or not, it represents a new endemic species to the assessment region. The species has still not been formally described (P. Taylor, pers. comm. 2025).
Red List Status: EN – Endangered
Assessment Information
Assessors: Taylor, P.1, & da Silva, J.M.2
Reviewer: Russo, I.3
Institutions:1University of the Free State,2South African National Biodiversity Institute, 3Cardiff University
Previous Assessors & Reviewers: Taylor, P., Willows-Munro, S., Baxter, R., Monadjem, A. & Child, M.F.
Previous Contributors: Roxburgh, L., Avenant, N.L., Avery, M., MacFayden, D., Palmer, G. & Wilson, B.
Assessment Rationale
This is a newly described species, possibly endemic to the Soutpansberg and Volkberg region of Limpopo Province. Further research is needed to determine whether the type specimen from Zuurbron, Wakkerstroom District in Mpumalanga Province, which is described as M. tenuis, is congruent with M. cf. tenuis. Similarly, comparisons of dental, morphometric, and molecular characters are needed to determine whether specimens assigned to M. cafer from Mpumalanga and Eswatini should be renamed M. cf. tenuis. Whether the final name and geographical distribution ultimately changes or not, this is certainly a species endemic to the assessment region.
It is an Afromontane grassland specialist existing in fragmented and possibly degraded patches. The extent of occurrence is estimated as 5,345 km2 while the area of occupancy (AOO), using the remaining natural Afromontane grassland (2014) as a proxy for suitable habitat, is estimated as 537 km2. However, this represents the maximum available AOO and does not incorporate potentially unoccupied patches, of which there are likely to be many given the fragmented nature of grassland patches and the poor dispersal capabilities of the species. Thus, we think it is reasonable to assume that true AOO is < 500 km2. This species is threatened by continuing declines in habitat area and habitat quality caused primarily by human settlement expansion, commercial plantations and mining, as well as associated overgrazing, water abstraction and fuelwood extraction relating to these land-use changes. Continuing decline is inferred from land satellite image and land-cover analysis: between 2000 and 2013, there has been a 14.9% and 8.8% rate of urban and rural expansion in the Limpopo Province, respectively; and 20% of forest and woodland cover was lost between 1990 and 2006 in the Soutpansberg Mountains due to residential expansion and pine/eucalyptus plantations. The latest trends post-2013 need to be examined to determine if this trend is declining and what proportion of the range is affected. Furthermore, climate change is an emerging threat to this species, as modelling predictions reveal a decline in suitable habitat ranging from 35–41% between 1975 and 2050. Vetting existing museum records to more accurately delimit range distribution for this new species, particularly in the Mpumalanga Province and Eswatini, is of utmost importance. Once these data have been generated, this species should be reassessed. For now, we assume it is a Limpopo Province endemic and list as Endangered B2ab (ii,iii,iv) under a precautionary purview, due to its restricted and severely fragmented area of occupancy, and an inferred continuing decline in habitat and habitat quality from human settlement, industrial and agricultural expansion in Limpopo Province. Key interventions include protected area expansion of Afromontane grassland and forest habitats, including the creation of corridors between patches and across elevational gradients to facilitate gene flow, as well as the enforcement of regulations restricting disturbance to suitable habitats.
Red List Index
Red List Index: No change
Regional Distribution and occurrence
Geographic Range
Recent molecular and morphological evidence argued for reassignment of populations formerly classified as M. cafer from the northern escarpment of South Africa and the Soutpansberg Mountains of the Limpopo Province to M. cf. tenuis (Taylor et al. 2013), pending formal description. Thus, currently, M. cf. tenuis is endemic to the Limpopo Province. However, pending ongoing research, the range may extend southwards to the type locality of M.tenuis(Zuurbron, Wakkerstroom District, Mpumalanga Province), and perhaps into Eswatini. The previous IUCN Red List assessment showed only the type specimen locality (Baxter & Hutterer 2008). It is strange that no museum specimens have yet been assigned to M. tenuis from the northern Drakensberg region of the Mpumalanga Province (being assigned rather to M. varius or, perhaps erroneously, M. cafer), which separates the Limpopo population from the type locality. A more accurate geographical distribution will be revealed following an analysis of such museum specimens.
Whether M. cf. tenuis it is a tropical or temperate species is unclear. Although subpopulations occupy montane grassland and forest habitats (Taylor et al. 2013), they occur mostly north of the Tropic of Capricorn (in the Soutpansberg Mountains) and at lower altitudes (up to 1,750 m asl in the Soutpansberg, and 2,050 m asl in the Wolkberg), which is characterised by moderate temperatures; for example, mean monthly temperatures measured at 1,750 m in the Soutpansberg varied from –0.4°C to 29.6°C (Taylor et al. 2015).
Using confirmed M. cf. tenuis records, the extent of occurrence in the Limpopo Province is estimated to be 5,345 km2. Using a recent land cover dataset from 2014 (GeoTerraImage 2015), area of occupancy (AOO) was estimated as 537 km2 by using remaining natural Afromontane grassland as a proxy for suitable habitat (sensu Taylor et al. 2017). However, it must be noted that this does account for potentially unoccupied patches and thus represents maximum available grassland. Similarly, the amount of available forest habitat is 181 km2. Grassland and forest habitat together is 718 km2. Since specimens have been collected primarily from moist grasslands and sometimes forest edges (but not within forests), using Afromontane grassland as a proxy for AOO is probably more accurate. They are sympatric in some areas with the more widespread M. varius.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
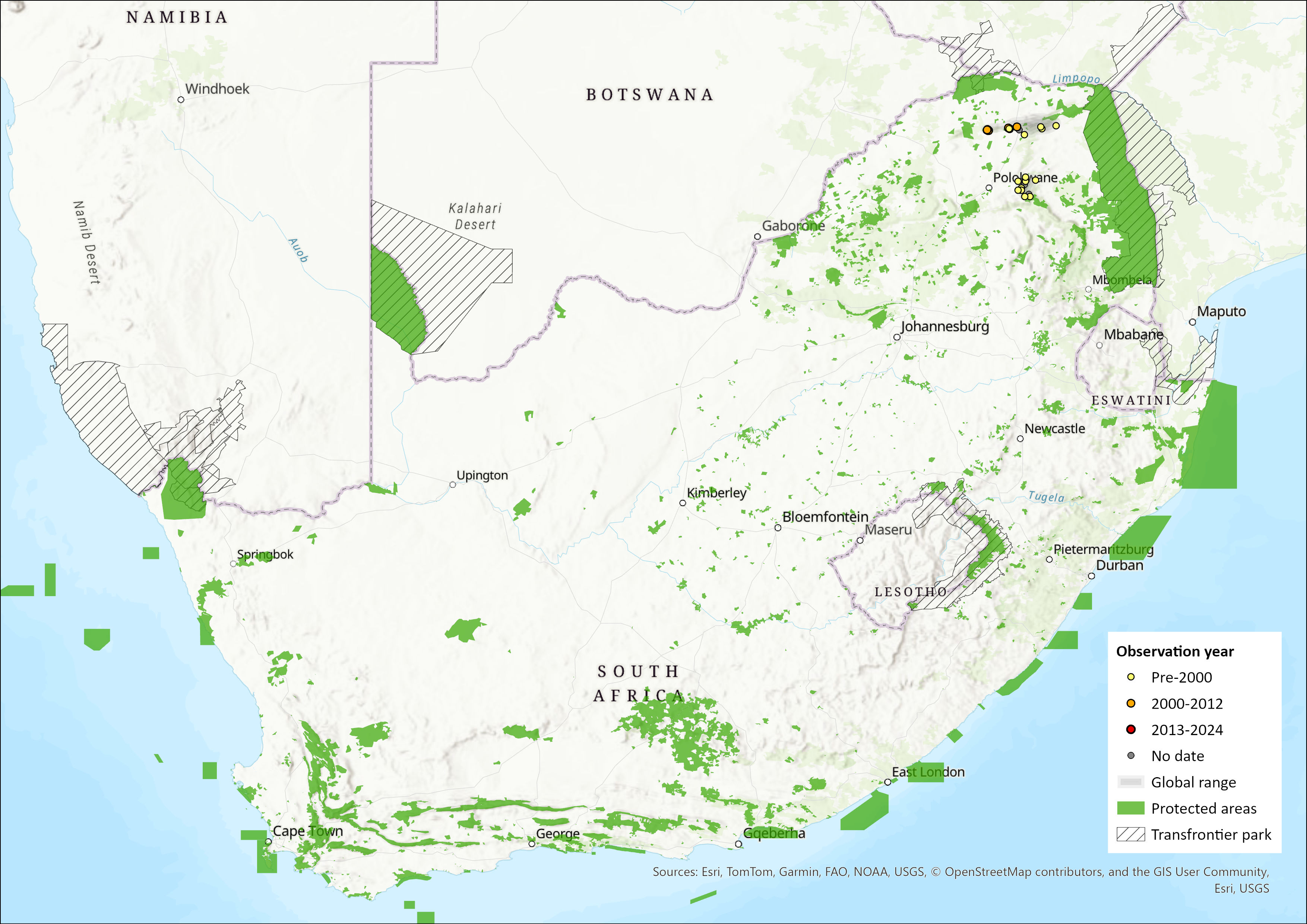
Figure 1. Distribution records for Thin Mouse Shrew (Myosorex tenuis) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin |
| Botswana | Absent | – |
| Lesotho | Absent | – |
| Mozambique | Absent | – |
| Namibia | Absent | – |
| South Africa: Limpopo | Extant | Native |
| South Africa: Mpumalanga | Presence uncertain | Native |
| Eswatini | Presence uncertain | Native |
| Zimbabwe | Absent | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Climate models project a 35-40% decline distribution of the species between 1975 and 2050 under both A1 and A2 scenarios (Taylor et al. 2017).
Population information
The abundance and population size of this species are not known. However, it may show an increase in abundance at higher elevations, similar to other Afromontane grassland small mammals (Taylor et al. 2015).
Population Information
Current population trend: Declining. Inferred from ongoing forest and grassland habitat loss.
Continuing decline in mature individuals: Unknown
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown, but perhaps as few as two, corresponding to the Soutpansberg and Wolkberg regions in the Limpopo Province.
Severely fragmented: Yes. Remaining Afromontane grassland and forest patches are patchily distributed between plantations, human settlements, mines and agricultural lands.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
Based on the mitochondrial control region and 16S rRNA markers, and the STAT nuclear marker (Taylor et al. 2013) nogenetic differentiation was found between the isolated northern Drakensberg and Soutpansberg populations. However, it is possible the markers were not robust enough to detect recent genetic differentiation. Considering populations east of the Sand River in the Soutpansberg are distinctly larger in cranial size than those to the west (Lajuma), while populations of Lajuma and the northern Drakensberg (Woodbush) are similar in size, it is very possible that genetically distinct subpopulations exist within the assessment region. It is highly recommended that more fine scale genomic markers be used to investigate the population genetic structure within this species.
Based on the available evidence it is expected that at least two genetic subpopulations exist. It is not possible to estimate effective population size (Ne) due to the lack of demographic population size estimates. In addition, the proportion of populations maintained (PM) indicator cannot be calculated with the current available data.
Habitats and ecology
The type specimen in Wakkerstroom, Mpumalanga Province was collected in thick grass at the edge of a stream at 1,600 m asl (Roberts 1951). In the Soutpansberg Mountains, Limpopo Province, they have always been collected from wetlands and moist grasslands, and some specimens were collected from the margins of mistbelt forests (Taylor et al. 2013), which emphasises the ecological separation between this species and the forest specialist M. cafer.
Ecosystem and cultural services: Candidate for flagship species in most grasslands biodiversity stewardship schemes in the Limpopo Province.
Life History
Generation Length: (Not specified)
Age at maturity: female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
There is no known subsistence or commercial use of this species.
General Use and Trade Information
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
The main threat to this species is the loss or degradation of moist, productive areas such as wetlands and rank grasslands in Afromontane habitats. The two main drivers behind this are habitat loss and abstraction of surface water and draining of wetlands through industrial (for example mining) and human settlement expansion, and overgrazing of moist grasslands, which leads to the loss of ground cover and decreases small mammal diversity and abundance (Bowland & Perrin 1989). Suppression of natural ecosystem processes, such as fire, can also lead to habitat degradation through bush encroachment or loss of plant diversity through alien invasives, and is suspected to be increasing with human settlement expansion. There are also clear overlaps and synergistic effects between these threats. Shrews have a high metabolic rate and thus rely on highly productive and complex environments, where small mammal diversity is highest (Bowland & Perrin 1993). Unfortunately, 65% of wetland ecosystem types within the assessment region are threatened (48% of all wetland types Critically Endangered, 12% Endangered and 5% Vulnerable; Driver et al. 2012).
Current habitat trend: Suitable montane grassland and woodland cover is declining within the Limpopo Province. There has been a significant expansion of the mining sector in recent years where prospecting and mining activities have been proposed or are currently underway in significant areas of high biodiversity, particularly in the Limpopo Valley, Soutpansberg, Blouberg, Makgabeng and Sekhukhune (V. Egan unpubl. data). Habitat loss and degradation is specifically apparent in the Soutpansberg, which is the core range of the species: significant habitat transformation occurred over the last 150 years (Hahn 2006), which is corroborated by recent land-cover analysis demonstrating that 20% of forest and woodland cover was lost from 1990 to 2006 in Soutpansberg Mountain region due to residential expansion and pine/eucalyptus plantations (Munyati & Kabanda 2009). Additionally, between 2000 and 2013, there was a 14.9% and 8.8% rate of urban and rural expansion in the Limpopo Province, respectively (GeoTerraImage 2015). This not only indicates loss of habitat outright, but increased pressure on ecosystems from human disturbance.
Conservation
There are no direct conservation measures in place for this species. It is not known if the species is present within any protected areas. Additional studies are needed into the taxonomy, distribution, abundance, general ecology and threats to this little-known species. However, we suspect the main intervention for this species is the protection and restoration of wetlands and grasslands. Protected area expansion should focus on connecting disparate patches of montane grasslands, which are highly fragmented (Neke & Du Plessis 2004). Biodiversity stewardship schemes should be promoted if landowners possess wetlands or grasslands close to core protected areas or remaining forest areas, and the effects on small mammal subpopulations should be monitored. Protecting such habitats may create dispersal corridors between suitable habitat patches.
Legislation should be enforced to prevent development or human encroachment in key habitats, which includes stricter zonation on development to decrease fragmentation of remaining grasslands.
At the local scale, landowners and managers should be educated, encouraged and incentivised to conserve the habitats on which shrews and small mammals depend. Retaining ground cover is the most important management tool to increase small mammal diversity and abundance. This can be achieved through lowering grazing pressure (Bowland & Perrin 1989), or by maintaining a buffer strip of natural vegetation around wetlands (Driver et al. 2012). Research will be needed to set the recommended length of the buffer strip in various habitats, but 500 m may provide a good indication of ecological integrity (Driver et al. 2012). Small mammal diversity and abundance is also higher in more complex or heterogeneous landscapes, where periodic burning is an important tool to achieve this (Bowland & Perrin 1993).
Similarly, the specific fire regime thresholds should be calibrated by research. Removing alien vegetation from watersheds, watercourses and wetlands is also an important intervention to improve flow and water quality, and thus habitat quality for shrews. This can be achieved through the Working for Water Programme (for example, Marais et al. 2004). However, the subsequent effects on shrew subpopulations must be monitored to demonstrate success (sensu Richardson & van Wilgen 2004). Education and awareness campaigns should be employed to teach landowners and local communities about the importance of conserving wetlands and moist grasslands.
Recommendations for land managers and practitioners:
- More accurate estimates of grassland patch occupancy through extensive live-trapping and field surveys should be conducted through dedicated surveys by specialists and conservation authorities to more accurately establish geographical range and potential biodiversity stewardship sites, thus informing spatial conservation planning.
- Enforce regulations on developments that potentially impact on the habitat integrity of grasslands and wetlands.
- Landowners should be incentivised to stock livestock or wildlife at ecological carrying capacity and to maintain a buffer of natural vegetation around wetlands.
Research priorities:
- Further vetting and analysis of museum specimens is needed to correctly identify and delimit the distributions of M. cafer and M. cf. tenuis, and molecular/morphological comparisons are needed to determine whether M. tenuis is the correct name for this species.
- Research should be conducted to determine disturbance thresholds in various habitats (for example, ecological stocking rates, amount of natural vegetation needed to sustain a viable subpopulation, and fire intensity and frequency needed to sustain habitat complexity) needed by managers to conserve shrew species.
Encouraged citizen actions:
- Citizens are requested to submit any shrews killed by cats or drowned in pools to a museum or a provincial conservation authority for identification, thereby enhancing our knowledge of shrew distributions (carcasses can be placed in a ziplock bag and frozen with the locality recorded).
Bibliography
Baxter R, Hutterer R. 2008. Myosorex tenuis. The IUCN Red List of Threatened Species 2008: e.T14115A4393755.
Bowland AE, Perrin MR. 1989. The effect of overgrazing on the small mammals in Umfolozi Game Reserve. Mammalian Biology 54:251–260.
Bowland JM, Perrin MR. 1993. Wetlands as reservoirs of small-mammal populations in the Natal Drakensberg. South African Journal of Wildlife Research 23:39–43.
Driver A, Sink KJ, Nel JN, Holness S, van Niekerk L, Daniels F, Jonas Z, Majiedt PA, Harris L, Maze K. 2012. National Biodiversity Assessment 2011: An Assessment of South Africa’s Biodiversity and Ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Friedmann Y, Daly B, editors. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. CBSG Southern Africa, IUCN SSC Conservation Breeding Specialist Group, Endangered Wildlife Trust, South Africa.
GeoTerraImage. 2015. Quantifying settlement and built-up land use change in South Africa.
Hahn N. 2006. Floristic diversity of the Soutpansberg, Limpopo Province, South Africa. Ph.D. Thesis. University of Pretoria, Pretoria, South Africa.
Jenkins PD, Churchfield S. 2013. Myosorextenuis Thin Mouse Shrew (Transvaal Forest Shrew). Pages 160–161 in Happold M, Happold D, editors. Mammals of Africa, Volume IV: Hedgehogs, Shrews and Bats. Bloomsbury Publishing, London, UK.
Marais C, van Wilgen BW, Stevens D. 2004. The clearing of invasive alien plants in South Africa: a preliminary assessment of costs and progress: working for water. South African Journal of Science 100:97–103.
Meester JA, Rautenbach IL, Dippenaar NJ, Baker CM. 1986. Classification of southern African mammals.Transvaal Museum Monographs 5:1–359.
Munyati C, Kabanda TA. 2009. Using multitemporal Landsat TM imagery to establish land use pressure induced trends in forest and woodland cover in sections of the Soutpansberg Mountains of Venda region, Limpopo Province, South Africa. Regional Environmental Change 9:41–56.
Neke KS, Du Plessis MA. 2004. The threat of transformation:quantifying the vulnerability of grasslands in South Africa. Conservation Biology 18:466–477.
Ogony OL. 2014. Potential impacts of climate change on Mysorex species as a model for extinction risk of montane small mammals in South Africa. M.Sc.Thesis. University of Venda, Thoyandou, South Africa.
Richardson DM,van Wilgen BW. 2004. Invasive alien plants in South Africa: how well do we understand the ecological impacts? South African Journal of Science 100:45–52.
Roberts A. 1951.The Mammals of South Africa. The Trustees of the Mammals of South Africa, Central News Agency, Johannesburg, South Africa.
Skinner JD, Chimimba CT. 2005. The Mammals of the Southern African Subregion. Third edition. Cambridge University Press, Cambridge, UK.
Taylor PJ, Kearney TC, Peterhans K, Julian C, Baxter RM, Willows- Munro S. 2013. Cryptic diversity in forest shrews of the genus Myosorex from southern Africa, with the description of a new species and comments on Myosorex tenuis. Zoological Journal of the Linnean Society 169:881–902.
Taylor PJ, Munyai A, Gaigher I, Baxter R. 2015. Afromontane small mammals do not follow the hump-shaped rule: altitudinal variation in the Soutpansberg Mountains, South Africa. Journal of Tropical Ecology 31:37–48.
Taylor PJ, Ogony L, Ogola J, Baxter RM. 2017. South African mouse shrews (Myosorex) feel the heat: using species distribution models (SDMs) and IUCN Red List criteria to flag extinction risks due to climate change. Mammal Research:1–14.
Willows-Munro S. 2008. The molecular evolution of African shrews (family Soricidae). Ph.D. Thesis. University of Stellenbosch, Stellenbosch, South Africa.