

Springhare
Pedetes capensis

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Pedetes capensis – (Forster, 1778)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – PEDETIDAE – Pedetes – capensis
Common Names: Springhare, South African Springhare (English), Springhaas (English and Afrikaans), Umahelane (isiNdebele), Tshipjane (Sesotho), Ntlolê, Ntlolane, Matlolane, Tshipo, Matsipane (Setswana), Nhire, Gwidzu (Shona), Ndlulwane (siSwati), Khadzimutavha (Venda), Indulane, Isandlulane (Zulu)
Synonyms: Yerbua capensis Forster, 1778, Pedetes albaniensis, Roberts 1946; Pedetes angolae, Hinton 1920; Pedetes cafer, Pallas 1778; Pedetes damarensis, Roberts 1926; Pedetes fouriei Roberts 1926; Pedetes orangiae, Wroughton 1907; Pedetes salinae, Wroughton 1907; Pedetes typicus, A Smith 1834.
Taxonomic Note: This species has not undergone any taxonomic changes since the 2004 assessment (Friedmann & Daly 2004) but the following key points are worth noting:
- The Pedetidae exhibit both hystricomorphous (porcupine-like) and sciurognathous (rat-like) characters but phylogenetic analyses support the grouping of the Pedetidae with the Sciurognathi (Matthee & Robinson 1997a).
- For a long time the genus Pedetes was thought to be represented by a single species, Pedetes capensis (Misonne 1972; de Graaf 1981; Meester et al. 1986; Dieterlin 1993), but it is now accepted (Skinner & Chimimba 2005) that there are two distinct species in this genus, P. surdaster from Kenya and Tanzania, and P. capensis from southern Africa. The recognition of the two species is based on phenotypical (Davies 1982), behavioural (Butynski 1978; Anderson 1996), placental (Otiang’a-Owiti et al. 1992), phylogeographical and cytogenetical differences between these species (Matthee & Robinson 1997b).
- Currently no subspecies of P. capensis are recognised within the assessment region (Meester et al. 1986; Matthee & Robinson 1997a; Skinner & Chimimba 2005).
Red List Status: LC – Least Concern
Assessment Information
Assessors: Oosthuizen, M.K.1, da Silva, J.M.2 & Russo, I.M.3
Reviewer: Kraai, M.4
Institutions: 1University of Pretoria,2South African National Biodiversity Institute,3Cardiff University,4Sol Plaatjie University
Previous Assessors and Reviewers: Peinke, D., Wilson, B. & Anderson, C.
Previous Contributors: Relton, C., Child, M.F., Avenant, N., Avery, M., Baxter, R., MacFadyen, D., Mondajem, A., Palmer, G. & Taylor, P.
Assessment Rationale
Springhares are widespread and abundant within the assessment region, occurring in various protected areas across their range. They utilise, and often prefer, cultivated and overgrazed environments. As such, there are no immediate threats to this species and there is no evidence of any obvious net population decline or range reduction. However, on a local scale, Springhares are particularly vulnerable to floods and persecution or overhunting. During country-wide surveys, a number of farmers reported localised extinctions and drastically reduced numbers following heavy past persecution. Whilst localised recolonisation over time is likely when culling efforts are ceased, the species provides an essential and mutualistic service to several other species (for example, Black-footed Cat, Felis nigripes (Brindley et al. 2024)) and is also an extremely important prey item for a range of species. Even a short period without the burrow refuge systems made available by Springhares could result in a devastating loss of safe burrow systems particularly in areas where other shelter options are limited. Thus, local extinctions of this species should be monitored as it may indicate broader biodiversity loss.
Regional population effects: There is presumably dispersal across the northern border of South Africa, especially across the contiguous arid habitat of the Kalahari. Immigration, and thus a rescue effect, is therefore possible but there is no evidence or reason to believe that there is currently a net movement of animals into or out of the country.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended Citation: Oosthuizen MK, da Silva JM & Russo IM. 2025. A conservation assessment of Pedetes capensis. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The species occurs across large parts of South Africa, Namibia, Botswana, Zimbabwe, Angola and Zambia as well as in the southern parts of the Democratic Republic of Congo and Mozambique (Skinner & Chimimba 2005). It is also predicted to occur marginally in the lowlands of Lesotho (Lynch 1994) and is listed in a recent compiled checklist of the mammals of Lesotho (Ambrose & Talukdar 2000). However, Boshoff and Kerley (2013) doubt the accuracy of these predictions and, until voucher specimens have been procured, their presence in Lesotho should be considered doubtful. Similarly, Butynski (2013) reports no records for either Lesotho or Eswatini.
Within South Africa, the species occurs across large parts of the Northern Cape, North West, Free State, Gauteng, Limpopo and Eastern Cape provinces, where there are sufficient sandy areas for burrowing, and open short grassland for foraging (Skinner & Chimimba 2005; Power 2014). In addition, it occurs in the western parts of Mpumalanga, the eastern areas of the Western Cape and marginally in the extreme northern and western parts of KwaZulu-Natal. It is absent from the eastern half of the Eastern Cape Province.
It is, however, important to note that within these areas their distribution is patchy and discontinuous. There is no clear evidence in the literature to indicate that the current distribution range has changed substantially from the historical distribution range. In Coetzee’s (1979) assessment of the distribution and status of some of the mammals of the Albany district (Eastern Cape Province). he does indicate that the species is absent from some areas where one might otherwise expect it to occur, and that this could be due to sport hunting. Although this seems to be localised and relatively insignificant, this trend throughout its range bears monitoring, as localised extinctions of subpopulations may be on the increase.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
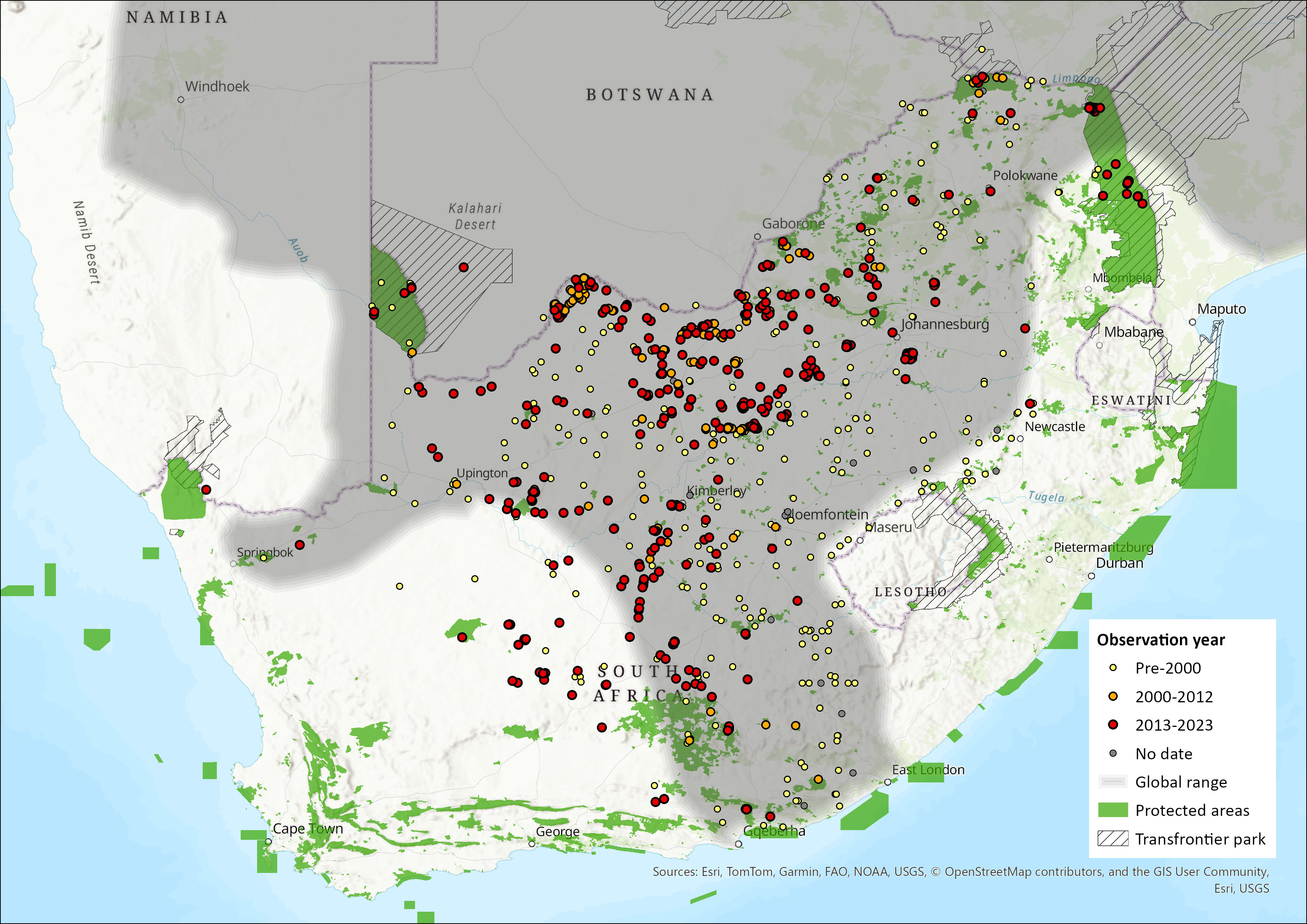
Figure 1. Distribution records for Springhare (Pedetes capensis) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Lesotho | Possibly Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
This species has not been specifically evaluated in terms of climate change. However, modelling shows a consistent habitat loss in the Nama Karoo for this species (Cowan 2025), and this appears to be the stronghold for this species. Springhare is widely distributed across the assessment range, and they are adapted to arid areas. Aridification would likely not affect the range of this species or affect it slower compared to other species. Burrowers tend to be less vulnerable and more protected against increasing temperatures compared to surface dwelling animals (Riddell et al. 2021). Nevertheless, changes in temperature and precipitation patterns can alter the habitats of burrowing species and affect the availability of food and water (Vale & Brito, 2015). Increases in extreme weather events can also negatively impact on animals, in all climates.
Population Information
This species is common to abundant across most of its range and is the most frequently encountered mammal while spotlighting in the more arid parts of the country (Power 2014). Highest densities are reached on flood plains and fossil lake beds (pans) where the vegetation is open, the grass is short and green, and sandy soils are available (Butynski 2013b). Population size is difficult to estimate but the following Springhare densities have been reported in the literature:
- Across seven sites in the Kimberley region, density ranged from 3 individuals / km2 in Kalahari Sandveld to 19 individuals / km2 in panveld (Anderson 1996); and 5 individuals / km2 on Benfontein Game Farm (Stenkewitz et al. 2010).
- Across fifteen sites in the Eastern Cape Province, densities ranged from 1 individual / km2 to 99 individuals / km2 (Peinke 2000; Peinke & Brown 2006).
- At Sandveld Nature Reserve, Free State Province: 40 individuals / km2 (Watson 1992).
- At SA Lombard Nature Reserve, North West Province: 10 individuals / km2 (van der Walt 1989).
Assuming that Springhare only occur over 30% of their defined area of occupancy (due to their need for deep sandy soils resulting in the patchy distribution) it can be estimated that there are conservatively between 2.23 million (8 x 278,700 km2) and 11.15 million (40 x 278,700 km2) Springhare in the country. Secondly, assuming that on average 74% of the aboveground population are adults (Butynski 1978; Anderson 1996; Peinke & Bernard 2005) then there are an estimated 1.65 and 8.25 million mature individuals in the national population. This is, however, only a very rough estimate and should be treated with extreme caution.
When only one Springhare species was recognized, it was listed as Vulnerable by the IUCN in 1996 due to an approximately 20% decrease in the population over the previous ten years. This was caused by intense hunting and the loss of habitat. This negative trend has not persisted, however, and the species is now listed as Least Concern. No current information is available on population trends, but the overall trend is suspected to be stable. Certain localised areas in the central Karoo, Eastern Cape and lower Kalahari have already reported lower numbers than those recorded 30–40 years ago. This may be indicative of a gradual decline as a result of persecution and severe competition with livestock such as sheep for forage. Additionally, this trend could also be attributed to climatic changes, associated with wetter conditions, which lead to habitat alteration in the form of taller grasslands. Long-term monitoring is needed to validate these observations and to quantify the decline, if it exists.
Population Information
Current population trend: Stable
Continuing decline in mature individuals: Unknown
Number of mature individuals in population: Approximately 1.65–8.25 million; although this is a vague estimate and should be treated with extreme caution.
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: No
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified) Not established
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified) Not established
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
While no population genetic studies have been conducted on this species, information can be inferred from the available population and distribution data to quantify the two proxy genetic indicators within the Convention on Biological Diversity’s Global Biodiversity Framework.
Because of its wide distribution and dispersal capabilities, it is thought this species exists as a metapopulation even connecting with neighbouring countries. Consequently, the GBF’s complementary genetic indicator – proportion of populations maintained (PM) – would receive a value of 1.0 (all populations remaining). While no census (Nc) or effective population size (Ne) data exists for the species, density estimates based on density estimates and AOO estimated that over 1.5 million Springhare exist within the assessment region (see Population section). Applying a 0.1-0.3 conversion ratio of these values, we get a possible Ne of over 150 000. From this GBF’s headline genetic indicator – proportion of populations with an Ne > 500 – is valued at 1.0 (1/1 population with Ne > 500). This population estimate far exceeds the Ne 500 threshold.
It is highly recommended that a population genetic study be conducted across the assessment region to verify the population genetic structure and diversity within this species.
Habitats and ecology
Springhares are the largest, hopping rodent species in southern Africa. With long back legs and short forelegs, they are kangaroo-like in appearance. With their long hind legs, they can reach speeds of 8.5 km / hour and jump distances of 0.8 m which is essential to avoid predators, a large number of which have been recorded utilising this species as prey. Despite their name, Springhares are neither hares nor rabbits but are rodents and are placed in their own unique genus. They have a sandy, cinnamon-coloured pelt, with a long bushy tail ending in a dark brown or black tip. Whilst there is some size variation regionally, the only significant colour variation recorded previously is from a very dark, almost melanistic, individual in the collection of the American Museum of Natural History (Butynski 2013b). In 2010, a similar specimen, almost jet-black, was collected near Cradock in the Eastern Cape and is now housed in the McGregor Museum collection (B. Wilson unpubl. data). Springhare fur is biofluorescent, with regions important for grooming and intraspecific interactions reflecting the most biofluorescence (Olsen et al. 2021). Biofluorescence is thought to have ecological significance for crepuscular or nocturnal mammals, and there is evidence of ecological tuning of biofluorescent signals in other species (Whicher et al. 2024).
With the exclusion of deserts and forested areas, the Springhare has a wide distribution in flat, arid and semi-arid areas. Pans and pan fringes can be considered optimum habitat. They prefer relatively flat and open habitats with short grass (in particular Cynodon spp.; Monadjem et al. 2015), usually where there is little or no woody vegetation (Smithers 1971; Butynski & Mattingly 1979; Butynski 1984, 2013b; Augustine et al. 1995; Anderson 1996; Skinner & Chimimba 2005). They are generally absent from areas of tall grass, dense woodland and Mopane (Colophospermum mopane) or miombo (Brachystegia spp.) where the soils are clay and poorly drained (Butynski 2013b). The species is a prolific burrower, remaining in any one burrow for only a few consecutive days (Peinke & Brown 2005) and, as such, is largely dependent on the presence of deep, compact, sandy soils that are suitable for burrowing into, and is absent from areas where the substrate is harder or unsuitable for burrowing (Smithers 1971; Butynski & Mattingly 1979; Coetzee 1979; Anderson 1996; Skinner & Chimimba 2005; Peinke & Brown 2006). Springhare also do well in areas that have been disturbed by cultivation and/or heavy grazing (Smithers 1971; de Graaf 1981; Augustine et al. 1995; Skinner & Chimimba 2005; Peinke & Brown 2006; Butynski 2013b). They are grazers feeding mainly on green grass seeds which are high in protein and water, but will also selectively feed on grass stems, leaves, corms, roots and rhizomes. As highly selective feeders, Springhare often uproot entire plants, feed on only chosen sections and discard the remainder (Skinner & Chimimba 2005).
The species is almost exclusively nocturnal but is sometimes encountered in the late afternoons during winter months. They forage in groups of two to six individuals however, group adhesion is not persistent, and members may join and leave with little reaction from others (Butynski 1984). Activity patterns are noticeably influenced by moonlight intensity, where they are known to forage much further from burrows (up to 400 m) during new moon cycles and remain closer to burrows (about 30 m) at periods of full moon (Anderson 1996). Springhares are non-territorial (Anderson 1996). Passive scent marking by means of a perianal gland takes place during feeding episodes, causing scent highways that the animal uses to locate its burrow quickly (Anderson 1996). This can be applied inside the burrow as well. Springhares are non-seasonal breeders, giving birth to one young after a gestation of an average of 77 days, and thus has a slow reproductive rate for a rodent (Butynski 1979; van der Merwe et al. 1980; Monadjem et al. 2015). Females may reproduce up to three times per year. Fatherhood is ensured by the depositing of a sperm plug into the female, thus preventing other males from impregnating that specific female (Anderson 1996). Its home range varies greatly between different individuals from 0.3 ha up to 28 ha (Peinke & Brown 2005). Burrows can cover an area of 170 m2, displacing approximately one ton of subsoil in the process (Anderson 1996). Burrows are occupied by a single animal or a mother and kitten.
Ecosystem and cultural services: This species is a keystone species in many of the ecosystems in which it occurs (Butynski 2013b). As such, the long-term survival of the species needs to be ensured. Firstly, Springhare burrows provide shelter and core areas for foraging for at least 20 other mammal, three bird, six reptile and 22 invertebrate species (Anderson 1996; Skinner & Chimimba, 2005). These include other threatened species such as Temminck’s Ground Pangolin (Smutsia temminckii) and Black-footed Cat (B. Wilson unpubl. Data; Brindley et al. 2024). The use of Springhare burrows by other species is particularly common in semi-arid regions of the Northern Cape, where burrows provide stable micro-environments away from extreme ranges in temperature and humidity (Skinnner & Chimimba 2005).
Additionally, Springhares are an important prey component within the food chains of all ecosystems in which they occur. From various studies on the species, Anderson (1996) and Skinner and Chimimba (2005) reported at least 20 mammals, seven birds and four reptiles as having preyed upon Springhares. One highly interesting case involved active hunting of Springhare by a pair of Bateleur Eagles (Terathopius ecaudatus) in the Kalahari, and this probably took place during late afternoon or moonlit nights (Anderson & Maritz 1994).
IUCN Habitats Classification Scheme
|
Habitat |
Season |
Suitability |
Major Importance? |
|
2.1. Savanna -> Savanna – Dry |
– |
Suitable |
– |
|
3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry |
– |
Suitable |
– |
|
4.5. Grassland -> Grassland – Subtropical/Tropical Dry |
– |
Suitable |
– |
|
14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land |
– |
Suitable |
– |
Life History
Generation Length: (Not specified)
Age at maturity: female or unspecified: 1034 days for females
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: 13-14 years in captivity
Average Reproductive Age: 8 months
Maximum Size (in cms): 80 cm
Size at Birth (in cms): 250-300g
Gestation Time: 77 days
Reproductive Periodicity: Breed throughout the year
Average Annual Fecundity or Litter Size: single young, up to 3 litters per year
Natural Mortality: Important prey species for at least 21 predator species (Butynski 1979)
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: No, typically a solitary species
Systems
System: Terrestrial
General Use and Trade Information
At a subsistence scale, Springhares are believed to be an important source of protein in rural communities. Butynski (1973) estimated that in Botswana 2.5 million Springhares are cropped annually for food. In 1991, the Board on Science and Technology for International Development National Research Council (BOSTID 1991) reported that at least 3.3 million kilograms of Springhare meat reached the Botswana market as the main source of bushmeat for human consumption in the country. To put this into perspective, this is equivalent to 30,000 cattle. Similar studies have yet to be conducted in South Africa and it is reasonable to assume that this practise extends into the assessment region too. The monitoring of subpopulations should be a priority in areas such as the Kalahari, and in North West and Limpopo provinces.
Recreational hunting, as a rite-of-passage for many young South African boys, is also known to take place on farms across its range. In commercial trophy hunting operations, Springhares are often offered as complimentary, non-trophy fee animals to clients. These may even enter the taxidermy industry afterwards for trophy mounting (B. Wilson unpubl. data).
There is limited commercial utilisation but where there is, it is assumed to be from the wild. However, there seems to be a growing market for the unusual and internet searches readily reveal prices such as US$150 for a skull, US$150 for a live animal as a pet, US$280–450 for a daily rate to hunt an animal, US$20–150 for trophy fees, and in game auctions in South Africa, between R60 and R400 for restocking purposes. There are even some websites providing information on how to raise and train them as pets.
Wildlife ranching has generally contributed positively to the conservation of the species by expanding and protecting available habitat. On game ranches Springhare are now often seen as an important game viewing species that contributes to visitor experience. This is quite different to the situation on commercial farms where Springhare are often regarded as pests because of damage to crops and competition with livestock.
Finally, apart from being a food source, the San have other cultural uses for Springhare. Skins are softened and used to make containers for food and water, as well as mats and karosses, while thread is derived from the sinews in the tail. Dung may even be mixed with other ingredients to make smoking tobacco.
|
Subsistence: |
Rationale: |
Local Commercial: |
Further detail including information on economic value if available: |
|
Yes |
– |
– |
– |
National Commercial Value: No
International Commercial Value: No
|
End Use |
Subsistence |
National |
International |
Other (please specify) |
|
1. Food – human |
true |
– |
– |
– |
|
10. Wearing apparel, accessories |
true |
– |
– |
– |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
While there are no quantified major threats to this species, its slow reproductive rate and sensitivity to disturbance means that several minor threats may cause local declines or extinctions:
- Persecution because of damage caused to crops. Butynski (1973) estimated that 10–15% of maize, sorghum, beans and groundnuts grown in Botswana were destroyed by Springhare but there is no information available on the extent of this problem or on the control measures implemented by farmers. The impact is felt on commercial crops, but probably more significantly by subsistence crop growers. Coetzee (1979) does report that Springhares are absent from some farms where one would expect them to be and indicates that this could be due to persecution. Their digging activities have also reportedly damaged roads, and anecdotal claims that eight foraging Springhares consume the same amount as a sheep are commonly heard from farmers. For these reasons, attempts to eradicate or significantly reduce resident subpopulations on some properties have resulted in localised declines and even extinctions in some cases. The general absence of information on this, however, suggests that it may not be a serious problem.
- Hunting for recreation and subsistence purposes (Butynski 1973; Coetzee 1979). Hunting for recreational and subsistence purposes takes place but there is no quantitative information available from the assessment region. As both recreational and subsistence hunting have been happening for many years without any apparent impact this may not be a serious problem. It is, however, important to note that Springhare are very easily hunted at night with a spotlight and consequently are very easily eradicated from areas. This, combined with a relatively low reproductive output (especially for a rodent), means that they could be vulnerable to high levels of hunting or utilisation.
- Habitat loss and transformation (Driver et al. 2012). Overall, 18% of South Africa’s surface area has already been irreversibly transformed mostly through cultivation, mining, forestry and urban development. In some regions the percentage of natural habitat lost is much higher and the rates of loss are alarming.
- Periods of abnormally high rainfall and floods will certainly affect Springhare subpopulation numbers, due to drowning. Additionally, persistently high rainfall levels, associated with climate change, may result in habitat loss, due to alteration in the form of increased grass height and woody cover. As a result of their fossorial habits, as well as their preference to colonise pan fringes (optimum habitat), Anderson (1996) modelled that excessive rainfall episodes is the biggest driving force in population regulation in the Northern Cape because of drowning.
This genus already has two extinct South African species of Pedetes known from fossil records (Butynski 2013), namely P. gracilis (Davies 1982) and P. hagenstadi (Cooke 1955), having occurred in the region of Taung, North West and Hagenstad Salt Pan (now known as Florisbad, Free State), respectively. These extinct species perhaps suggest the genus is vulnerable to disturbance and provide a cautionary tale for current conservation.
Habitat trend: Stable. The species occupies a wide variety of habitats, including agricultural landscapes. However, the loss of grassy areas or pans will impact local subpopulations.
Conservation
As a keystone species, the long-term survival of the species needs to be ensured. Springhares are well conserved in several protected areas scattered across their distribution range, including Augrabies Falls National Park, Kgalagadi Transfrontier Park, Tswalu Game Reserve, Molopo Nature Reserve and the Mountain Zebra National Park. Conservationists must ensure that these protected areas are adequately funded and managed. No specific conservation interventions are necessary at present. However, long-term monitoring schemes and research into the capacity for sustainable harvest of this species should be initiated (see Management Recommendations and Research Priorities). However, hunting should be regulated in areas where the species is declining.
Recommendations for land managers and practitioners: Currently there are no conservation plans for the species, and the creation of such plans is undermined by a lack of data of subpopulation health and trends. It can be reasonably expected that the numbers will decline in some regions and localised areas. These areas need to be identified.
Persecution as a problem-causing species will continue, but judicious and sustainable harvesting techniques must be employed to minimise the impact on the populations.
- Initiate long-term monitoring programmes to provide information on population trends.
- Review legislation related to the hunting and harvesting of this species.
- Establish sustainable harvesting guidelines, considering the slow breeding rate.
- Develop management plans in areas where numbers have significantly dropped.
Research priorities:
- The economic value of the species.
- Levels of sustainable harvest, under various natural and man-made conditions, and the extent to which Springhares are hunted for recreational and subsistence reasons in South Africa.
- The effect of environmental conditions and population density on behaviour.
- Quantification and description of agricultural damage caused by Springhares on a national scale, how they are managed in problem areas and if this constitutes a significant threat to the species.
- The degree of commensalism with this species and others (particularly protected or threatened species) to determine its importance in the ecosystem as a refuge provider.
- Comparative ecological, behavioural and physiological studies with the equatorial P. surdaster.
- The status and population trends associated with subpopulations.
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Establish citizen science monitoring programmes. Springhare are very easy to monitor and it would be useful if some long-term population monitoring sites could be established on public and private land across the range of this species. This information could be coordinated at provincial level and fed into regional databases used for decision-making during the conservation ordinance amendment processes. Ideally spotlight counts should be conducted along set routes and repeated on at least three consecutive nights. Preferably, data should be collected during the first half of the evening, on moonless nights (Peinke & Brown 2006) and during mid-winter and again in summer. For even better density estimates, the distance sampling method, described in Stenkewitz et al. (2010) could be implemented; however, this would require additional effort from the monitors. Data should be submitted to provincial coordinators, and subsequently to the national coordinator. Should citizens be willing to establish long-term monitoring sites, the assessment authors may be contacted.
Bibliography
Ambrose D, Talukdar S. 2000. Biological Diversity in Lesotho. National Environment Secretariat, Maseru, Lesotho.
Anderson PC, Maritz AWA. 1994. Prey items of two pairs of Bateleur Eagles in the Northern Cape Province, South Africa. Gabar 9: 26-27.
Anderson PC. 1996. The population dynamics and ecological role of the springhare Pedetes capensis (Forster, 1778) in the Kimberly area, Nothern Cape Province, South Africa. Ph.D. Thesis. University of the Free State.
Augustine, D. J., Manzon, A., Klopp, C. and Elter, J. 1995. Habitat selection and group foraging of the springhare, Pedetes capensis larvalis Hollister, in East Africa. African Journal of Ecology 33: 347-35.
BOSTID. 1991. Microlivestock: Little-known Small Animals with a Promising Economic Future. National Academy of Sciences, National Academy Press, Washington, DC, USA.
Boshoff AF, Kerley GIH. 2013. Historical incidence of the larger mammals in the Free State Province (South Africa) and Lesotho. Centre for African Conservation Ecology, Nelson Mandela Metropolitan University, Port Elizabeth.
Brindley, H., Justin O’Riain, M. and Sliwa, A. 2024. The underground cat: burrow use by female black-footed cats (Felis nigripes). African Zoology 1-12.
Butynski TM, Mattingly R. 1979. Burrow structure and fossorial ecology of the springhare Pedetes capensis in Botswana. African Journal of Ecology 17: 205–215.
Butynski TM. 1979. Reproductive ecology of the springhaas Pedetes capensis in Botswana. Journal of Zoology 189: 221–232.
Butynski TM. 1984. Nocturnal ecology of the springhare, Pedetes capensis, in Botswana. African Journal of Ecology 22: 7–22.
Butynski TM. 2013a. Family Pedetidae. In: Happold DCD (ed.), Mammals of Africa, Bloomsbury Publishing, London, UK.
Butynski, T.M. 1973. Life history and economic value of the springhare (Pedetes capensis Forster) in Botswana. Botswana Notes and Records 5: 200-2013.
Butynski, T.M. 2013b. Pedetes capensis Southern African Springhare. In: D.C.D. Happold (ed.), Mammals of Africa. Volume III: Rodents, Hares and Rabbits, Bloomsbury Publishing, London.
Coetzee, P. W. 1979. Present distribution and status of some of the mammals of Albany. Albany Divisional Council and Grahamstown Municipality, South Africa.
Cooke HBS. 1955. Some fossil mammals in the South African Museum collections. Annals of the South African Museum 42: 161–168.
Cowan, OS. 2025. Modelling the change in potential species distribution of Springhare (Pedetes capensis) under predicted future climate change scenarios. Technical Report, Endangered Wildlife Trust.
Davies C. 1982. The recent and fossil affinities of the genus Pedetes (Mammalia: Rodentia). Ph.D. Thesis. University of Oxford.
De Graaff, G. 1981. The Rodents of Southern Africa. Butterworths, Durban, South Africa.
Dieterlin F. 1993. Family Pedetidae. In: Wilson DE, Reeder DM (ed.), Mammal Species of the World: a Taxonomic and Geographic Reference, pp. 1207. Smithsonian Institution Press, Washington, DC, USA.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., Van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An assessment of South Africa’s biodiversity and ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Friedmann, Y. and Daly, B. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. Conservation Breeding Specialist Group (SSC/IUCN) and Endangered Wildlife Trust, Parkview, South Africa.
Lynch, C.D. 1994. The mammals of Lesotho. Navorsinge van die Nasionale Museum Bloemfontein 10(4): 177-241.
Matthee CA, Robinson TJ. 1997a. Molecular phylogeny of the springhare, Pedetes capensis, based on mitochondrial DNA sequences. Molecular Biology and Evolution 14: 20–29.
Matthee, C.A. and Robinson, T.J. 1997b. Mitochondrial DNA phylogeography and comparative cytogenetics of the springhare, Pedetes capensis (Mammalia: Rodentia). Journal of Mammalian Evolution 4: 53-73.
Meester, J.A.J., Rautenbach, I.L., Dippenaar, N.J. and Baker, C.M. 1986. Classification of Southern African Mammals. Monograph number 5. Transvaal Museum, Pretoria, South Africa.
Misonne X. 1972. Order Rodentia. In: Meester J, Setzer H (ed.), The Mammals of Africa: An Identification Manual., pp. 1–39. Smithsonian Institution Press, Washington, DC, USA.
Monadjem, A., Taylor, P.J., Denys, C. and Cotterill, F.P.D. 2015. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis. De Gruyter, Berlin, Germany.
Olson, E.R., Carlson, M.R., Ramanujam, V.M.S., Sears, L., Anthony, S.E., Anich, P.S., Ramon, L., Hulstrand, A., Jurewicz, M., Gunnelson, A.S., Kohler, A.M., Martin, J.G. 2021. Vivid biofluorescence discovered in the nocturnal springhare (Pedetidae). Scientific Reports 11:4125.
Otiang’a-Owiti GE, Oduor-Okelo D, Gombe SG. 1992. Foetal membranes and placenta of the springhare (Pedetes capensis larvalis Hollister). African Journal of Ecology 30: 74–86.
Peinke D, Bernard RTF. 2005. Life history of the springhare (Pedetes capensis) from a strongly seasonal environment in the Eastern Cape Province of South Africa. African Zoology 40: 285–292.
Peinke DM, Brown CR. 2005. Burrow utilization by springhares (Pedetes capensis) in the Eastern Cape, South Africa. African Zoology 40: 37–44.
Peinke DM, Brown CR. 2006. Habitat use by the southern springhare (Pedetes capensis) in the Eastern Cape Province, South Africa. South African Journal of Wildlife Research 36: 103–111.
Peinke DM. 2000. The Ecology and Physiology of the springhare (Pedetes capensis) in the Eastern Cape Province of South Africa. Ph.D. Thesis. Rhodes University.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Riddell, E.A., Iknayan, K.J., Hargrove, L., Tremor, S., Patton, J.L., Ramirez, R., Wold, B.O., Beissinger, S.R. 2021. Exposure to climate change drives stability or collapse of desert mammal and bird communities. Science 371:6333-636.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Smithers, R.H.N. 1971. The mammals of Botswana. National Museums of Rhodesia, Museum Memoir 4: 1-340.
Stenkewitz U, Herrmann E, Kamler JF. 2010. Distance sampling for estimating springhare, cape hare and steenbok densities in South Africa. South African Journal of Wildlife Research 40: 87-92.
Vale, C.G., Brito, J.C. 2015. Desert-adapted species are vulnerable to climate change: Insights from the warmest region on earth. Global Ecology and Conservation 4:369-379.
van Der Merwe M, Skinner JD, Millar RP. 1980. Annual reproductive pattern in the springhaas, Pedetes capensis. Journal of Reproduction and Fertility 58: 259–266.
Watson J. 1992. Beraamde springhaas populasie van die Sandveld Natuurreservaat. Free State Nature and Environmental Conservation, South Africa.
Whitcher, C., Ron, S.R., Ayala-Varela, F., Crawford, A.J., Herrera-Alva, V., Castillo-Urbina, E.F., Grazziotin, F., Bowman, R.M., Lemmon, A.R., Lemmon, E.M. 2024. Evidence for ecological tuning of anuran biofluorescent signals. Nature Communications 15:8884.