

springbok
Antidorcas marsupialis

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Antidorcas marsupialis – (Zimmermann, 1780)
ANIMALIA – CHORDATA – MAMMALIA – ARTIODACTYLA – BOVIDAE – Antidorcas – marsupialis
Common Names: Springbok (Afrikaans and English), Springbuck (English), Tshephe (Sotho), Ibhadi (Xhosa), Insephe (Zulu)
Synonyms: No Synonyms
Taxonomic Note:
Meester et al. (1986) listed three subspecies: A. m. marsupialis from the southern part of the range; A. m. hofmeyri from Botswana, Namibia and the Northern Cape; and A. m. angolensis from Angola. While the subspecies distinction is debated, morphometric data reveal a difference in size between Springbok occurring on either side of the Orange and Vaal Rivers, which is evidence for maintaining subspecies status (Peters & Brink 1992). Further taxonomic research is thus needed to determine the status of two distinct ecotypes from the north (Kalahari region) and the south (Karoo region).
Red List Status: LC – Least Concern
Assessment Information
Assessors: Patel, T.1; da Silva, J. M.2
Reviewer: Mallon, D.3
Institutions: 1Endangered Wildlife Trust; 2South African National Biodiversity Institute; 3IUCN SSC Antelope Specialist Group
Previous Assessors: Anderson, C., Schulze, E., Codron, D. & Anderson, J.
Previous Reviewers: Child, M.F., Bissett, C. & Gaylard, A.
Previous Contributors: Selier, J.
Assessment Rationale
Springbok are listed as Least Concern due to their widespread distribution within the assessment region, and an estimated mature population size of 76,445 – 77,545 animals (2016 assessment). However, local or regional declines have occurred, most notable of which is the subpopulation in Karoo National Park, which declined by 63% between 1994 and 2015 (2,163 to 794 animals). This was precipitated by a large cull in 2000 with a continuing decline thereafter. The most recent data received for Karoo National Park (2019) did not include population numbers. The causes of such declines are unclear but may involve environmental stresses (such as extended wet periods) and degradation (from excessive livestock overgrazing), allee effects, mesopredator pressure and poaching (localised contexts) – the relevance or severity of such threats varying by area. Overall, imprudent translocation of locally adapted ecotypes (Kalahari versus Karoo) and the emerging threat of selective breeding for rare colour variants (where the “regular” Springbok ewes are simply treated as surrogate uteri) may cause maladaptive traits, which could prove detrimental to the species in the face of climate change. Such threats should be managed through improving connectivity between habitats and regions to allow for greater dispersal and gene flow, as well as the development of a scientifically informed national translocation policy. It is also recommended that this species could be used sustainably for game-meat production as part of wildlife-based rural economies, where subpopulations are performing adequately. Predicted habitat changes due to climate change could increase the importance of Springbok in producing protein off arid rangelands. This species is a national symbol of South Africa and must continue to be conserved as an abundant and resilient population.
Regional population effects: There is dispersal across the South African, Namibian and Botswanan borders, especially within Kgalagadi and Richtersveld Transfrontier Parks. There are very few other free-roaming subpopulations in the assessment region. The majority of the population is kept on private land within the natural distribution of the species.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Patel T & da Silva JM. 2025. A conservation assessment of Antidorcas marsupialis. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The Springbok’s distribution is mainly confined to southern Africa, except for a narrow extension into southwestern Angola, where it inhabits the South West Arid Zone and adjacent dry savanna of south and southwestern Africa. It still occurs very widely within its historical range, but in Angola it survives in greatly reduced numbers (East 1999, Skinner 2013). It does not occur in Eswatini and became Extinct in Lesotho from overhunting (Lynch 1994).
In South Africa, the Springbok was almost exterminated over much of its natural range in the Free State, the former Transvaal and, to a large extent, in the former Cape Province, during the 19th century as a result of hunting, rinderpest and farm fences blocking their migratory routes. However, the species has subsequently been reintroduced widely to private land and protected areas throughout its former range and beyond. The largest numbers occur on private game farms, mainly in the Highveld of the Free State and Gauteng provinces, as well as the Karoo and Kalahari thornveld of the Western, Eastern and Northern Cape provinces. In the North West Province, they occur throughout the southern and western arid and open areas, where a free-roaming herd occurs, and may be expanding, between the western Kalahari, near Terra Firma, and the Highveld grasslands further southeast (Power 2014). They thus currently occur in all provinces, especially in larger protected areas and many private lands in central and western South Africa. The majority of subpopulations held on private land are still within the natural distribution of the species. However, smaller, introduced populations occur widely in extralimital areas, such as on private land in parts of KwaZulu-Natal and the northern bushveld (Mpumalanga and Limpopo provinces) (East 1999). Such introductions have made it difficult to determine the natural distribution range of the species (Skinner and Louw 1996, Cain et al. 2004).
Feely (2014) documented historical records recorded by the English artist, Thomas Baines, for Springbok occurring in the northeastern part of the Eastern Cape in 1848 (on the edge of the natural range). There was previously a relict population of the former eastern Transvaal Springbok population on the farm Blair Athol near Amsterdam surviving since it was first surveyed in the late 1800s (J. Anderson, pers. comm. 2014). There are, unfortunately, no Springbok left here (J. Anderson, pers. comm. 2025).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
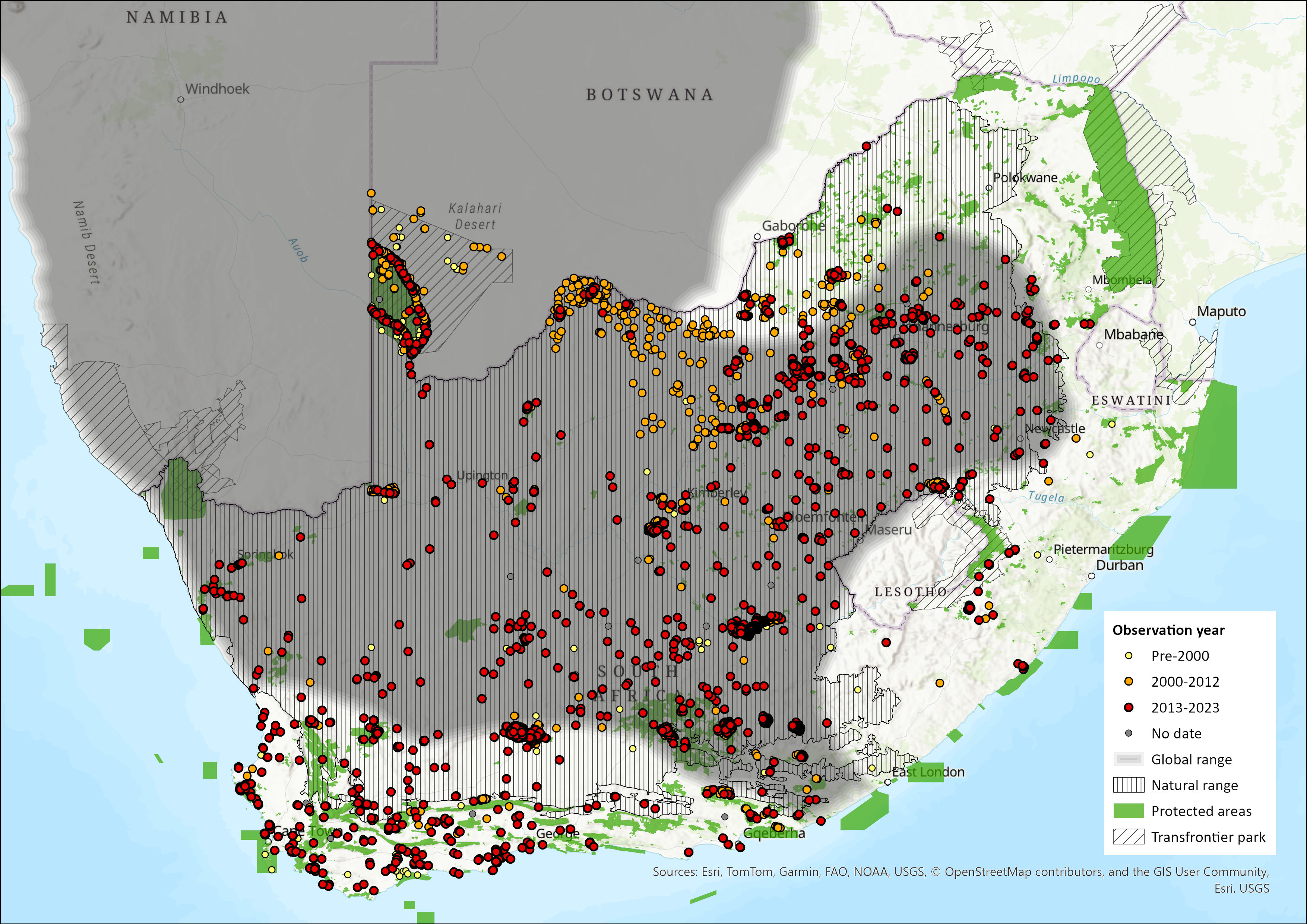
Figure 1. Distribution records for Springbok (Antidorcas marsupialis) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Lesotho | Extinct Post-1500 | Native | – | – |
| Namibia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Springbok subpopulations vary in relation to climatic conditions (Skinner and Chimimba 2005). In a study conducted in the Kalahari Gemsbok National Park, it was found that springbok behavior, particularly feeding and food selection, was significantly affected by environmental and seasonal conditions. This implies that changes in climatic conditions, as predicted by climate change, can negatively impact springbok behavior (Stapelberg et al. 2008). In addition, selective breeding for rare colour variants can result in maladaptive traits which impacts their ability to adapt to changing environments in the face of climate change.
Population
Recent data captured from provinces and from the private sector were not robust enough to update the population numbers from the 2016 assessment, which were based on 654 reserves and properties. There is no evidence to suggest a change in population size that would affect the conservation status of this species at present. The population numbers cited are based on what we consider wild individuals.
Skinner (2013) estimated the total population size for Springbok in southern Africa at c. 2,000,000–2,500,000 animals, with approximately 60% of the population occurring on privately-owned land (East 1999). For provinces where fairly comprehensive estimates of the private Springbok population are available (Free State and North West), we estimate a higher proportion of the population on private land: 76–89%. Specifically for South Africa, Skinner (2013) estimated 75,000 in the Free State, 75,000 in the former Transvaal provinces, 1,000,000 in the Karoo and about 100,000 in the Cape provinces outside of the Karoo. However, these are most likely overestimates based on observed game count data in South Africa between 2013 and 2015. For example, it was estimated to be only 42,731 animals in Free State and 109,209–110,778 in total across the country. As the private sector data was incomplete in most provinces, this figure was likely to be an underestimate. Given that the proportion of mature individuals in a herd is typically 70–73% (C. Anderson unpubl. data), we infer a total mature population of at least 76,446–77,545 animals, based on the 2013-2015 counts. The largest subpopulation occurs on the South African side of Kgalagadi Transfrontier Park, where Springbok numbers were 1,935 (wet season count) and 2,217 (dry season count) in 2012 (Ferreira et al. 2016). Recent data were received for the Kgalagadi in 2019; however, records did not include population counts. While the number of cross-border Springbok dropped dramatically in the late 1980s, they have stabilised since with no dramatic declines being recorded in recent years. It is unclear why there was a dramatic decline. It is possible that human-induced factors such as fences, roads and cattle post development were responsible for this decline (Mills and Mills 2013).
Chelsmford Nature Reserve in KwaZulu-Natal falls within the natural distribution range and thus is included in the population estimate. While no formally protected subpopulations exist in Limpopo, some private subpopulations do. It is unclear whether these are extralimital as property coordinates are not available. They are included in the population estimate. Similarly, for Mpumalanga, while one private nature reserve is known to fall within the natural range, the other is not (due to missing coordinates). For formally protected areas, while we include Nooitgedachtdam Dam Nature Reserve (as it is on the border of the natural range) we exclude Songimvelo Nature Reserve as it falls significantly outside the natural range. In both Limpopo and Mpumalanga provinces (as well as the Cape provinces), the numbers of private subpopulations are likely to be significantly more extensive, but no comprehensive data are available at present. Further collation of such data is important. However, not all private subpopulations will count towards the wild population if they are intensively managed. Further research is required to determine which private subpopulations are eligible.
Generation length has been estimated as seven years (Pacifici et al. 2013), making the three-generation window 21 years (1994–2015). Based on 30 formally protected areas from across the natural range of Springbok with adequate long-term data available, we estimated a net 8–27% national increase in population size over three generations (14,927 to 16,542 animals). It is well-known that Springbok experience population fluctuations. Thus, further long-term datasets should be made available to estimate population trend more accurately. There are also worrying examples of local subpopulation declines across its range, the largest of which has been seen in Karoo National Park, having declined from 2,163 to 794 animals between 1994 and 2015 (Gaylard et al. 2016). Reasons for the decline are largely unknown but may include Allee effects following a crash after intense culling in 2000 (of c. 50% of the subpopulation) and subsequent park expansion. The most recent data received for Karoo National Park reflected 766 individuals (SANParks data; 2021 count). Subpopulations also show mixed success in the North West Province. For example, at Bloemhof Dam Nature Reserve, numbers have increased from 479 in 1999 to 1,151 in 2023, but have declined in Molopo Nature Reserve recently by 14% between 2014 and 2015, contrary to expected as the veld was dominated by short grass especially around the pans (Nel 2015). This trend is still being seen in Molopo from 2021 to 2023, where the population declined from 233 individuals to 189 individuals. They have declined especially in Mafikeng Nature Reserve from 738 in 1999 to 108 in 2015. They have thus declined even further to 90 individuals in 2020 and 71 in 2021.
Subpopulation declines (since 2009) have also been recorded in parts of the Free State (E. Schulze unpubl. data), North West (Nel 2015) (especially on communal lands, Buijs 2010) and Northern Cape (M. Smit unpubl. data) provinces, possibly due to predation, habitat changes and disease. For example, the decline in numbers in the Kimberley area from 2009 until 2013 (5,573 to 4,855 animals), excluding the sharp spike in 2010 (6,826 animals), can be directly attributed to the higher than normal rainfall experienced during this period (C. Anderson unpubl. data), where most of the pans, which are prime habitat and feeding spots for Springbok, were inundated with water and resulted in taller grass stands. It is well known that a wet substrate and tall grass stands are not optimal habitat for Springbok (Skinner and Chimimba 2005). The occurrence of “hoof-rot” (vrotpootjie) was also reported on certain properties (C. Anderson unpubl. data). A rapid turnaround of this situation took place from 2014 into 2015, when the area experienced a severe dry spell, and a rapid improvement in the rate of recruitment, as well as more synchronised lambing, was experienced. Springbok in the Northern Cape normally show distinct lambing peaks in autumn (April–May) and spring (September). This was completely disrupted during the excessively wet period, which also increased the predator (jackal) impact on the herds. Another phenomenon experienced during periods of stress were the birth of more males than females (Krüger et al. 2005), as well as physical signs (limited) of foetal resorption (tiny foetuses) (examination of foetuses by C. Anderson).
Population Information
Current population trend: Stable
Continuing decline in mature individuals? Local and regional declines from predation, habitat changes and disease but inferred to be stable overall.
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: No
Number of mature individuals in population: At least 76,446–77,545
Number of mature individuals in largest subpopulation: South African side of the Kgalagadi Transfrontier Park: 1,034–4,332 individuals (Ferreira et al. 2016).
Severely fragmented: Yes. Most subpopulations occur in fenced protected areas or ranch lands. However, the major subpopulation occurs as a free-roaming subpopulation in the Kgalagadi Transfrontier Park.
Number of subpopulations: 655 sites
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, 7maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
While a comprehensive population genomic study has not been undertaken on this species across its range, several molecular studies looking at the of genetic diversity within and among different springbok colour variants have been (Kruger 1976, Skead 2007, Bezuidenhout 2012, Coetzer & Grobler 2019). These studies have been motivated by concerns that intensive human-mediated line-breeding for rare colour morphs (such as black, white, or copper springbok) may lead to a reduction in genetic variability. However, comparable levels of genetic diversity were observed for all colour variants tested, suggesting no loss of diversity during line breeding (Coetzer & Grobler 2019).
With respect to genetic structure, most colour variants showed no distinct genetic differentiation. An exception was the white springbok, which exhibited slight genotypic divergence and formed a separate cluster (Coetzer & Grobler 2019). This could reflect both historical cultural and hunting practices (white springbok were revered and not hunted by indigenous peoples, but selectively targeted by early colonial hunters; Roche, 2005), as well as modern artificial selection, which together may have led to a degree of genetic drift within this phenotype (Coetzer & Grobler 2019).
It is important to note that these colour variants are typically intensively managed and largely confined to private or commercial game farms. As such, they are generally considered ineligible when assessing natural population trends. Despite this, some colour morphs, including white springbok, are occasionally observed in free-roaming populations of common springbok (Hetem et al. 2009), suggesting some level of historical or incidental gene flow. Nonetheless, these instances appear to be rare, and wild populations are likely largely insulated from these artificially maintained variants. Accordingly, only a single population (metapopulation) is recognised for this species within the assessment region, based on natural free-roaming populations and excluding artificially maintained colour variants.
Based on the recognition of a single metapopulation and approximately 76,445 – 77,545 individuals across the country, the effective population size (Ne) for the species in the assessment region is projected to exceed the Ne 500 threshold (assuming a Ne/Nc ratio of 0.1-0.3: Ne ~7 600-23 400).
Habitats and ecology
The Springbok is the most abundant plains antelope in the arid lands of southern Africa and formerly occurred in huge numbers in the dry grasslands and shrublands of southwestern and southern Africa, migrating sporadically in vast herds (“trekbokken”) in some of the southern parts of its range in response to rainfall and the search for verdant veld (Skinner 1993). For example, Cronwright-Schreiner (1925) observed a herd of hundreds of thousands attempting to cross the Orange River where thousands of animals drowned. These migrations or treks no longer occur, having largely been eliminated by expansion of stock farming, and associated fencing, overhunting and the rinderpest epidemic in 1896 (Skinner 1993). However, some indication of the species’ former abundance can still be seen as seasonal concentrations in areas with preferred short vegetation in parts of the Kalahari that occur in central and southern Botswana (East 1999). Congregations disperse into small groups during winter (Cain et al. 2004).
Springbok are mixed feeders (Hofmann et al. 1995), consuming both browse and grass. They prefer to utilise grass when it is young but otherwise browse karroid vegetation which includes a variety of low shrubs and succulents (Skinner 2013). Nevertheless, they show great variation in dietary preferences across habitats, and even amongst individuals within habitats (D. Codron pers. obs.), as some individuals and even whole subpopulations consume mainly grass. Springbok occur in areas where surface water is unavailable or available only seasonally and receive their moisture requirements from browsing on succulent karroid vegetation, by digging succulent roots, or by eating fruits such as Solanum spp. (Nagy and Knight 1994, Skinner and Chimimba 2005). They are often sympatric with merino sheep in the Karoo, where Springbok display a better spatial utilisation of the vegetation by being more dispersed in the landscape and less dependent on proximity of food to water points that prevent over-exploitation of the vegetation (Davies and Skinner 1986a).
Within the matrix of Kalahari sands, Springbok prefer both salt and calcareous pans (Milton et al. 1992), as well as dry riverbeds (Jackson 1997), and the males select territories to attract the attention of female herds by choosing these high-quality foraging habitats (Jackson and Skinner 1998). These high-quality habitats tend to have more clay soils (aside from dry riverbeds and Kalahari sands) that are nutrient rich and have a richer assemblage of dwarf shrubs. Such habitats are, however, localised in occurrence, and are represented by the Southern Kalahari Mekgacha and Southern Kalahari Salt Pans vegetation (Mucina and Rutherford 2006). The presence of such habitat types may thus influence Springbok spatial ecology (Power 2014). Key habitats are thus open grasslands (particularly the short grass fringes of pans), arid shrubveld (Karoo) and the mainly karroid vegetation associated with Northern Cape pans.
Ecosystem and cultural services: The Springbok is the national animal of South Africa. It also features prominently in San paintings demonstrating that they have been significant to humans for thousands of years (Eastwood et al. 2002)
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 4.5. Grassland -> Grassland – Subtropical/Tropical Dry | – | Suitable | – |
| 8.2. Desert -> Desert – Temperate | – | Marginal | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: Congregatory (and dispersive)
Systems
System: Terrestrial
.
General Use and Trade Information
Springbok are hunted and traded as live animals, as well as for their horns, meat and skins (IUCN SSC Antelope Specialist Group 2008). Ranching of Springbok for subsistence and commercial venison has been common since the early 1900s (Skinner and Louw 1996). Mixed herds of Springbok and Merino sheep (Ovis aries) are common farming combinations in the Karoo (Davies and Skinner 1986b). The percentage of animals taken from ranched stock is unknown. Protected areas, however, manage subpopulations carefully. For example, Free State provincial reserve managers harvest approximately 15–20% of the Springbok population annually (E. Schulze unpubl. data).
Trade is suspected to be leading to an increase in the population as more ranchers breed, sell and protect Springbok. However, the intensive selective breeding for colour variants should be discouraged as this may lead to inbreeding which affects the resilience of the overall population. Colour variants may also show maladaptive traits for increased temperature due to climate change. Hetem et al. (2009) compared the physiology of black, white and normal Springbok in the Karoo and found that the colour morphs exhibit differences in body temperature and activity consistent with differences in solar heat load: while black Springbok had lower diurnal activity in winter (having to forage less because their metabolic cost of homeothermy was lower), they were disadvantaged in hot periods. White Springbok, by contrast, were more protected from solar heat load (similar to that of the normal type in summer), but potentially less able to meet the energy cost of homeothermy in winter. Thus, energy considerations may underlie the rarity of natural Springbok colour morphs (Hetem et al. 2009). Maintaining genetic diversity, especially for a trait like pelage colour, rather than fixing certain traits, is thus of adaptive advantage to the Springbok population when responding to climate change (sensu Millien et al. 2006).
| Subsistence: | Rationale: | Local Commercial: | Further detail including information on economic value if available: |
| Yes | Species is used locally as a meat source. | Yes | Used nationally and internationally for meat, live sales and trophy hunts. |
National Commercial Value: Yes
International Commercial Value: Yes
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? Yes
Harvest Trend Comments: Extensive ranching occurs. The species is harvested for meat, trophy hunts, and live sales. A small proportion of subpopulations are harvested sustainably as part of hunting packages or to control herbivore numbers. The species is intensively bred for colour variants and sold to other breeders.
Threats
Within the assessment region, while the overall population is stable, Springbok numbers have declined in some areas, while increasing elsewhere due to trade. The possible causes of decline include environmental stress (wet conditions with tall grass stands), poaching, predation and disease. This decline is probably manageable, but multiple stakeholder involvement might be required. Springbok subpopulations vary in relation to climatic conditions (Skinner and Chimimba 2005), and sound land management is required to prevent degradation of pans and bush encroachment. Additionally, increased mesopredator abundance may impact some subpopulations (sensu Minnie et al. 2016). Poaching is a localised threat. For example, at Boskop Dam Nature Reserve in North West Province, poaching is suspected to be causing some decline due to the presence of snares (Nel 2015). Diseases may also cause subpopulation declines. For example, Springbok do not occur in woodland savannah due to the presence of Heartwater to which they show no resistance (Neitz 1944). Hybridisation between ecotypes may constitute a threat to some subpopulations (Friedmann and Daly 2004). Game ranchers may be mixing southern (Karoo region) and northern (Kalahari region) ecotypes for sport hunting (as the latter animals are larger, Mills and Mills 2013, L. Rossouw and J.S. Brink unpubl. data). This mixing could lead to outbreeding depression due to maladaptation of the Kalahari form to the rockier substrate of southern habitats (that may increase mortality with no noticeable effect on fecundity) and increase the prevalence of disease, such as “hoof-rot”. An emerging threat to this species is the intensive and selective breeding of the species within the wildlife industry (Bezuidenhout 2012). Artificially selecting individuals to produce rare colour variants may also cause maladaptive traits (for example, Hetem et al. 2009).
Conservation
Springbok are well represented in protected areas throughout their range, the largest being Kgalagadi Transfrontier Park between Botswana and South Africa. Springbok are also abundant in private lands in South Africa, where they are actively managed. Springbok are amongst the most valued species in the expanding game ranching industry in southern Africa due to the excellent quality of their venison (Hoffman et al. 2007, Skinner 2013). However, it is crucial that artificially selected individuals do not enter the national or provincial parks systems. Thus, appropriate legislation and a translocation and management plan should be developed and enforced to restrict movement from private ranches into formally protected areas (which is already in place for SANParks and many provincial conservation authorities). Land managers should also be incentivised to drop internal fences to form conservancies, which may allow Springbok subpopulations greater movement to conserved key habitat patches.
Recommendations for land managers and practitioners:
- Management and translocation plan to restrict movement and dispersal of privately-owned Springbok, which may contain hybridised individuals or colour variants into formally protected areas.
- Greater government subsidies should be for infrastructure to enhance the use of Springbok as a key species in building sustainable wildlife-based economies. Springbok meat is very lean with fat content not exceeding 4% (Skinner and Louw 1996). Optimal annual cropping rate is 30% of the animals; however, in years with high rainfall, cropping rate may be increased to 40% (Skinner and Louw 1996).
Research priorities:
- Research is required to determine the causes of local declines in populations in the Kgalagadi Transfrontier Park and other Provincial and National Parks.
- Undertake taxonomic research to determine the status of two suspected subspecies or ecotypes pertaining to the north (Kalahari) and south (Karoo).
- Assess the potential of Springbok to become a source of sustainable, low-carbon protein.
- Quantify the extent and impacts of hybrids and artificially-bred colour variants.
- Understand the impact of predators on the population dynamics of this species.
- Determine the effect of global warming on Springbok population dynamics.
- Determine the impact of mesopredators on the population dynamics of this species.
Encouraged citizen actions:
- Report sightings on virtual museum platforms (e.g., iNaturalist and MammalMAP) outside of protected areas.
- Create conservancies to sustain wild and free-roaming herds of Springbok and other species.
Bibliography
Bezuidenhout R. 2012. Colour under investigation. Farmer’s Weekly. Johannesburg, South Africa..
Buijs, D. 2010. Aerial survey of wildlife and domestic stock on tribal land in North West Province 2010. DACERD, Mmabatho.
Cain JW, Krausman PR, Germaine HL. 2004. Antidorcas marsupialis. Mammalian Species 753: 1-7.
Cronwright-Schreiner SC. 1925. The Migratory Springbucks of South Africa. Fisher Unwin, London, UK.
Davies RAG, Skinner JD. 1986a. Temporal activity patterns of springbok Antidorcas marsupialis and merino sheep Ovis aries during a Karoo drought. Transactions of the Royal Society of South Africa 46: 133-147.
Davies RAG, Skinner JD. 1986b. Spatial utilisation of an enclosed area of the Karoo by springbok Antidorcas marsupialis and Merino sheep Ovis aries during drought. Transactions of the Royal Society of South Africa 46: 115-132.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., Van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An assessment of South Africa’s biodiversity and ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
East, R. (compiler). 1999. African Antelope Database 1998. IUCN, Gland, Switzerland and Cambridge, UK.
Eastwood E, van Schalkwyk J, Smith B. 2002. Archaeological and rock art survey of the Makgabeng Plateau, Central Limpopo Basin. The Digging Stick 19: 1-3.
Feely JM. 2014. Historical incidence of springbok (Antidorcas marsupialis) in the northeastern Eastern Cape: further evidence. South African Journal of Wildlife Research 44: 71-75.
Ferreira S, Gaylard A, Greaver C, Hayes J, Cowell C, Bissett C. 2016. Large vertebrate abundances in Parks: 2015/2016. Internal Report 14/2016, Scientific Services, SANParks, Skukuza, South Africa.
Friedmann, Y. and Daly, B. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. Conservation Breeding Specialist Group (SSC/IUCN) and Endangered Wildlife Trust, Parkview, South Africa.
Gaylard A, Ferreira S, Bezuidenhout H, Smit I. 2016. Karoo National Park aerial census report 2015. SANParks Internal Report 05 / 2016, Scientific Services
Hetem RS, de Witt BA, Fick LG, Fuller A, Kerley GI, Meyer LC, Mitchell D, Maloney SK. 2009. Body temperature, thermoregulatory behaviour and pelt characteristics of three colour morphs of springbok (Antidorcas marsupialis). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology 152: 379-388.
Hoffman LC, Kroucamp M, Manley M. 2007. Meat quality characteristics of springbok (Antidorcas marsupialis). 4: Sensory meat evaluation as influenced by age, gender and production region. Meat Science 76: 774-778.
Hofmann RR, Knight MH, Skinner JD. 1995. On structural characteristics and morphophysiological adaptation of the springbok (Antidorcas marsupialis) digestive system. Transactions of the Royal Society of South Africa 50: 125-142.
IUCN SSC Antelope Specialist Group. 2008. Antidorcas marsupialis. The IUCN Red List of Threatened Species 2008: e.T1676A6359255. Available at: http://dx.doi.org/10.2305/IUCN.UK.2008.RLTS.T1676A6359255.en.
Jackson TP, Skinner JD. 1998. The role of territoriality in the mating system of the springbok Antidorcas marsupialis. Transactions of the Royal Society of South Africa 53(271-282).
Jackson TP. 1997. Springbok Antidorcas marsupialis. In: M.G.L. Mills and L. Hes (eds), The Complete Book of Southern African Mammals, pp. 262-263. Struik Publishers, Cape Town, South Africa.
Krüger O, Radford AN, Anderson C, Liversidge R. 2005. Successful sons or superior daughters: sex–ratio variation in springbok. Proceedings of the Royal Society of London B: Biological Sciences 272: 375-381.
Lynch, C.D. 1994. The mammals of Lesotho. Navorsinge van die Nasionale Museum Bloemfontein 10(4): 177-241.
Masubelele ML, Hoffman MT, Bond WJ, Gambiza J. 2014. A 50 year study shows grass cover has increased in shrublands of semi-arid South Africa. Journal of Arid Environments 104: 43-51.
Meester, J.A.J., Rautenbach, I.L., Dippenaar, N.J. and Baker, C.M. 1986. Classification of Southern African Mammals. Monograph number 5. Transvaal Museum , Pretoria, South Africa.
Millien V, Kathleen Lyons S, Olson L, Smith FA, Wilson AB, Yom-Tov Y. 2006. Ecotypic variation in the context of global climate change: revisiting the rules. Ecology letters 9: 853-869.
Mills G, Mills M. 2013. A Natural History Guide to the Arid Kalahari including the Kgalagadi Transfrontier Park. Black Eagle Media (Pty) Ltd, Cape Town, South Africa.
Milton SJ, Dean WRJ, Marincowitz CP. 1992. Preferential utilization of pans by springbok (Antidorcas marsupialis). Journal of the Grassland Society of Southern Africa 9: 114-118.
Minnie L, Gaylard A, Kerley GI. 2016. Compensatory life-history responses of a mesopredator may undermine carnivore management efforts. Journal of Applied Ecology 53: 379-387.
Mucina, L. and Rutherford, M.C. 2006. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria, South Africa.
Nagy KA, Knight MH. 1994. Energy, water, and food use by springbok antelope (Antidorcas marsupialis) in the Kalahari Desert. Journal of Mammalogy 75: 860-872.
Neitz, W. O. 1944. The susceptibility of the Springbuck (Antidorcas marsupialis) to heartwater. Onderstepoort Journal of Veterinary Science and Animal Industry 20: 25-27.
Nel, P. 2015. Population estimates for large herbivores and predators in protected areas in the North West Parks Board November 2015. North West Parks Board.
Pacifici, M., Santini, L., Di Marco, M., Baisero, D., Francucci, L., Grottolo Marasini, G., Visconti, P. and Rondinini, C. 2013. Generation length for mammals. Nature Conservation 5: 87–94.
Peters J, Brink JS. 1992. Comparative postcranial osteomorphology and osteometry of springbok, Antidorcas marsupialis (Zimmerman, 1780) and grey rhebok, Pelea capreolus (Forster, 1790) (Mammalia: Bovidae). Navorsinge van die Nasionale Museum Bloemfontein 8: 161-207.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Skinner JD, Louw GN. 1996. The springbok Antidorcas marsupialis (Zimmerman 1780). Transvaal Museum Monographs 10: 1-50.
Skinner JD. 1993. Springbok (Antidorcas marsupialis) treks. Transactions of the Royal Society of South Africa 48: 291-305.
Skinner, J. D. 2013. Antidorcas marsupialis. In: J. S. Kingdon and M. Hoffmann (eds), The Mammals of Africa, Academic Press, Amsterdam, The Netherlands.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Stapelberg H, Rooyen V, W M, Bothma J du P, Linde VD, J M, Groeneveld HT. 2008. Springbok behaviour as affected by environmental conditions in the Kalahari. Koedoe 50: 145-153.