

Southern Savannah Buffalo
Syncerus caffer caffer

2025 Red list status
Least Concern
Regional Population Trend
Increasing
Change compared
to 2016
No Change
Overview
Syncerus caffer caffer – (Sparrman, 1779)
ANIMALIA – CHORDATA – MAMMALIA – ARTIODACTYLA – BOVIDAE – Syncerus – caffer
Common Names: Southern Savannah Buffalo, Cape Buffalo, African Buffalo, Buffalo (English), Buffel, Afrikabuffel (Afrikaans), Inyathi (Ndebele, Xhosa, Zulu), Nare (Sepedi, Sesotho, Setswana), Inyatsi (Swati), Nari (Venda), Nyarhi (Tsonga)
Synonyms: No Synonyms
Taxonomic status: Subspecies
Taxonomic Note:
Four African Buffalo subspecies e.g. S. c. nanus, S. c. brachyceros, S. c. aequinoctialis and S. c. caffer (Michaux et al. 2023).The East and southern African, the group is comprised of a single subspecies (Syncerus caffer caffer), which is found in the assessment region (Michaux et al. 2023). The West and Central African group is assigned to three subspecies (S. c. nanus, S. c. brachyceros and S. c. aequinoctialis) (Le Roex et al. 2012, Smitz et al. 2013, Michaux et al. 2023).
The West African Savannah Buffalo, S. c. brachyceros (from Senegal to Cameroon) and the Central African Savannah Buffalo, S. c. aequinoctialis (from Central Africa to Sudan) are morphologically intermediate between S. c. caffer (Southern Savannah Buffalo) and S. c. nanus (Forest Buffalo) (Skinner and Chimimba 2005). Forest Buffalo are adapted to forest life, having a smaller size, unobtrusive swept back horns and a red to reddish-brown colouration, and are restricted to West Africa (Skinner and Chimimba 2005). Southern Savannah Buffalo are about twice the size of Forest Buffalo, with large downward curved horns and a brownish to black colouration (Smitz et al. 2013). The taxonomic uncertainty in these four subspecies is corroborated by the observation of interbreeding and intermediate phenotypes in contact zones between all four subspecies (Prins 1996), and at least one reported a cross between S. c. caffer and S. c. nanus (Smitz et al. 2013). Based on recent genetic results, S. c. brachyceros and S. c. aequinoctialis would be included in S. c. nanus following standard nomenclature rules (Smitz et al. 2013). Evidence suggests that buffalo populations in Uganda and the Central African Republic display the highest genetic diversity of all African Buffalo populations, and this could be the region where the most overlap exists between the two main groups (S. c. nanus and S. c. caffer; Smitz et al. 2013). Analysis of 766 mtDNA sequences from 43 localities indicated that S. c. caffer is found in Uganda, Kenya, Tanzania, Zambia, Angola, Zimbabwe, Botswana, South Africa and Namibia (Smitz et al. 2013).
| Red List Status |
| LC – Least Concern, (IUCN version 3.1) |
Assessment Information
Assessors: Venter, J.1, Russo, I.M.2 & da Silva, J. M.3
Reviewer: Anderson, J.4,5
Contributor: Patel, T.6
Institutions: 1Nelson Mandela University, 2Cardiff University, 3South African National Biodiversity Institute, 4IUCN SSC Antelope Specialist Group, 5International Conservation Services, 6Endangered Wildlife Trust
Previous Assessors: Venter, J. & Tambling, C.
Previous Reviewer: Child, M.F.
Previous Contributor: Du Toit, J.
Assessment Rationale
This subspecies is widespread and abundant within the assessment region, with subpopulations being reintroduced across the country on private lands both within and outside their original range (which should be regulated). The total mature population size in 2013 was estimated at 50,231 animals in 70 formally protected areas (35,162 mature animals), while the private sector is estimated to have added at least 19,561 further animals, raising the total population size to at least 69,882 animals (48,917 mature). However, many of these private subpopulations are intensively managed and thus not eligible for Red List assessments. Future assessments should determine the true wild and free-roaming population sizes. The national population is estimated to have significantly increased over three generations (1982–2015) and threats that could cause local subpopulation declines (such as disease, drought and poaching) are mitigated by the game ranching industry that has stimulated increasing numbers of reintroductions and introductions onto private properties. However, the increasing intensive and selective breeding of private subpopulations may be lowering genetic diversity and this should be monitored. Overall, while local declines must be monitored and managed, and genetic diversity should be sustained through a translocation policy, this subspecies should continue to thrive within the assessment region. Thus, the Least Concern listing remains.
Regional population effects: Although there will be some movement of buffalo between the Kruger National Park (KNP), Mozambique and Zimbabwe, it is likely that the KNP subpopulation will be providing a source for the Zimbabwe (possibly) and Mozambique sections of the Transfrontier Park. Fence barriers are negotiated with relative ease, particularly across rivers. A 2010 census of Limpopo National Park in Mozambique revealed that most buffalo sightings were in the southwestern sections and close to the KNP (Wildlife Management Services – Parque Nacionale do Limpopo 2010), suggesting movement from the KNP into Limpopo National Park. Buffalo have also dispersed onto most wildlife ranches adjoining the KNP in Mozambique.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Venter J, Russo IM & da Silva JM. 2025. A conservation assessment of Syncerus caffer caffer. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Please scroll horisontal on mobile device to see the full table
Regional Distribution and occurrence
Geographic Range
Before the influence of European settlers, the African Buffalo’s former range stretched over most of southern Africa and Angola, through central and east Africa to the southern borders of Sudan and Ethiopia (Sinclair 1977). The Southern Savannah Buffalo subspecies is distributed through the eastern and southern portions of the overall range, predominantly in Uganda, Kenya, Tanzania, Zimbabwe, Botswana and South Africa (Smitz et al. 2013); and is also still present in Namibia and Mozambique (Winterbach 1998). Disease has played a significant role in buffalo geographical population status that we see today (de Jager et al. 2021). Only about 5% of buffalo in the region survived the rinderpest and foot-and-mouth disease epidemics of the late 1890’s and early 1900’s (Winterbach, 1998). In the mid-1900s they were exposed to further extermination as part of the tsetse-fly controll program (Winterbach, 1998). The population recovered from three sources e.g. Kruger National Park, Hluhluwe-iMfolozi Park and Addo Elephant National Park (Michaux et al. 2023). Since then, the population expanded and are now generally confined to protected areas (fenced and unfenced), within which they are well represented.
The largest subpopulations of buffalo are in the savannah areas of the Limpopo Province, Mpumalanga and KwaZulu-Natal (KZN; Winterbach 1998), and to a lesser degree the Eastern Cape Province. They have been reintroduced to areas from which they were formerly extirpated (for example, in the North West Province; Power 2014). Similarly, they were reintroduced in Eswatini, where the indigenous population was extirpated. There are fewer populations in the Western and Northern Cape Provinces (Winterbach 1998; Venter 2006). Buffalo distribution in South Africa is expanding rapidly with buffalo ranching and breeding operations becoming more popular (Venter 2006).
It should be noted that in the stocking of Madikwe, in order to source disease-free animals, some buffalo from European zoos were brought in. These were thought to be from founders that originated from East Africa (Boonzaaier pers comm.).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 0 metres
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Biogeographic Realms
Biogeographic Realm: Afrotropical
Map
Click on map to view/enlarge
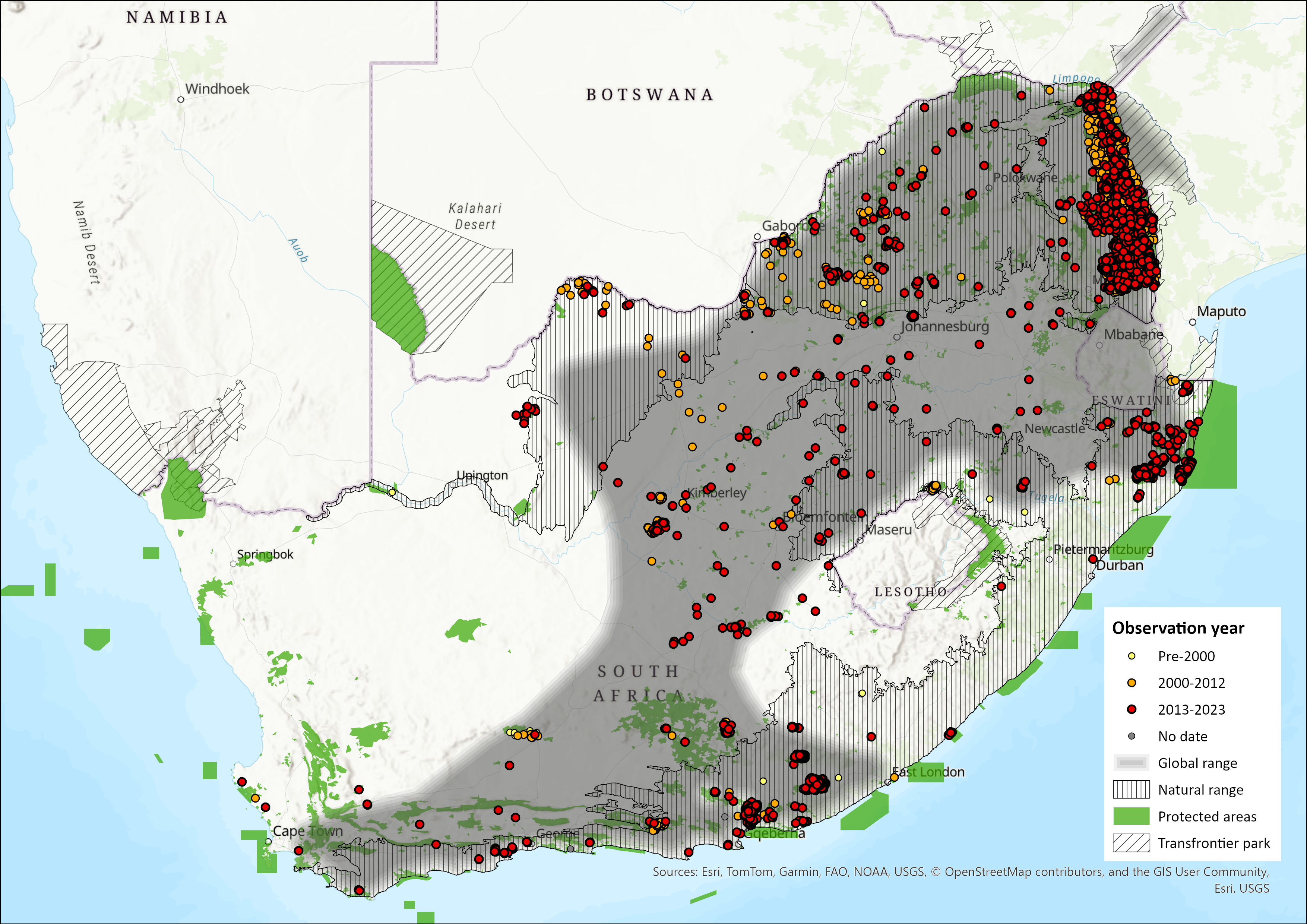
Figure 1. Distribution records for Southern Savannah Buffalo (Syncerus caffer caffer) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Countries of Occurrence
| Angola | Extant | Native | – | – |
| Benin | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Burkina Faso | Extant | Native | – | – |
| Burundi | Extant | Native | – | – |
| Cameroon | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Chad | Extant | Native | – | – |
| Congo | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Côte d’Ivoire | Extant | Native | – | – |
| Equatorial Guinea | Extant | Native | – | – |
| Eritrea | Extinct Post-1500 | Native | – | – |
| Eswatini | Extant | Reintroduced | – | – |
| Ethiopia | Extant | Native | – | – |
| Gabon | Extant | Native | – | – |
| Gambia | Extinct Post-1500 | Native | – | – |
| Ghana | Extant | Native | – | – |
| Guinea | Extant | Native | – | – |
| Guinea-Bissau | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Liberia | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mali | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Niger | Extant | Native | – | – |
| Nigeria | Extant | Native | – | – |
| Rwanda | Extant | Native | – | – |
| Senegal | Extant | Native | – | – |
| Sierra Leone | Extant | Native | – | – |
| Somalia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Togo | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
The Southern Savannah Buffalo is a water dependent species that requires on average about 30-40 liters of water per day (Du Toit 2005a). Animals prefer to utilise natural water sources while grazing preferentially on grasses close by these water sources (Du Toit 2002, 2005). Given this dependence on water, reductions in surface water availability would have negative impacts on the subspecies. Southern Savannah Buffalos are also sensitive to heat, so future increases in temperature regimes might have the potential to adversely affect the species (Heinz 2012).
They are also vulnerable to drought (Nel 2015), which historically resulted in significant population declines, especially when associated with diseases, such as rinderpest or anthrax; for example, during the 1990s, in Tsavo National Park (Kenya), the Serengeti/Mara (Tanzania), Gonarezhou National Park (Zimbabwe) and KNP (South Africa) (East 1999). Within the assessment region, drought is related to lack of forage availability after having been artificially provided (increasingly the case on private properties). The effects of climate change may exacerbate these problems, rendering many areas unsuitable for this subspecies.
Population information
It has been estimated that 670,000 Southern Savannah Buffalo exist across the continental distribution range (IUCN SSC Antelope Specialist Group 2008). In many parts of its range, the numbers of African Buffalo are thought to be declining because of bushmeat hunting and continuing loss of habitat (IUCN SSC Antelope Specialist Group 2008). Winterbach (1998) estimated that a population of 111,900 buffalo inhabit southern Africa (excluding Mozambique).
Within the assessment region, there were an estimated 50,231 animals in 70 formally protected areas (2013 counts), which yields a mature protected population of 35,162 animals (assuming a 70% mature population structure). There were at least an additional 19,561 animals on 110 private properties across the country (2013 counts). This yields a total population size of 69,882 animals (48,917 mature) in 2013. However, many private buffalo subpopulations are intensively managed (Taylor et al. 2015) and do not qualify for inclusion in the Red List assessment. Further work is thus required to determine the wild and free roaming population size. The largest subpopulation exists in KNP, estimated at over 27,000 (Michel et al. 2006) and, more recently, 37,322 animals (Ferreira et al. 2013). The KNP subpopulation fluctuates in response to rainfall and drought, with population crashes observed during droughts in the 1980s and 1990s (Funston & Mills 2006). Additionally, the KNP population has been infected with Bovine Tuberculosis since 1990 (de Vos et al. 2001), along with foot and mouth disease, corridor disease and brucellosis. The subpopulation in the Hluhluwe-iMfolozi Park in KZN is estimated at 5,468 animals (Clinning 2012) and the iSimangaliso Wetland Park contains around 1600 animals (Church pers comm. 2025). The Hluhluwe-iMfolozi population is infected with Bovine Tuberculosis and both these and the iSimangaliso animals are carriers of corridor disease. The largest subpopulations in the Eastern Cape Province are estimated to be 399 in Addo Elephant National Park (Ferreira et al. 2013) and 318 in the Great Fish River Nature Reserve (Peinke & Gibisela 2014; 2013 counts). The population in Madikwe (448) is also affected with Bovine Tuberculosis and that in Loskop Dam Nature Reserve (c. 200) with corridor disease (Theileriosis). The Doornkloof Nature Reserve in the Northern Cape Province has a population of c. 100 animals (2013 count). The efforts to produce disease-free buffalo from infected stock have been summarised by Laubscher & Hoffman (2012).
Generation length for this species has been estimated as 11 years (Pacifici et al. 2013), yielding a 33 three–generation window (1982–2015). Using 11 formally protected areas, with adequate long-term data over this time, we estimated a population increase of 398% over three generations (1,888 to 9,401 animals on the sampled protected areas), with only two sampled subpopulations experiencing declines over this period. Thus, the population in the assessment region is currently increasing whereas they may be declining elsewhere in Africa (IUCN SSC Antelope Specialist Group 2008; Craigie et al. 2010). While there are local fluctuations (such as in the KNP subpopulation) related to climatic variability (droughts), buffalo have proven to recover rapidly to former levels after such events.
Population Information
Current population trend: Increasing
Continuing decline in mature individuals: No
Number of mature individuals in population: 35,162 – 48,917
Number of mature individuals in largest subpopulation: 26,125
Number of subpopulations: 70 (formally protected areas)
Severely fragmented: Yes, populations are largely restricted to national parks and private lands, and dispersal is limited by fences.
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
Population genetic studies using microsatellites and mitochondrial DNA have indicated high levels of genetic differentiation between subspecies and between populations of the Southern Savannah Buffalo across Africa (O’Ryan et al. 1998; Van Hooft et al. 2000; Smitz et al. 2014; De Jager et al. 2020). Specifically, two evolutionarily significant units (ESUs) have been identified within the species, with only one in the assessment region (Smitz et al. 2013). Despite the two not being reciprocally monophyletic and the presence of shared haplotypes, ESU status is warranted given that the two have different chromosome numbers (S. c. caffer: 2n = 52 and S. c. nanus: 2n = 54), which may lead to reduced fitness (outbreeding depression) of hybrids, and there has been very low levels of gene flow between the two lineages in the recent past.
Additional substructure has been found within the S. c. caffer ESU of which three subpopulations exist in the assessment region. These include a subpopulation in north-eastern South Africa and Mokala NP (central South Africa, disease-free population of Kruger NP buffalo). The Hluhluwe-iMfolozi Park subpopulation (HiP), and the Addo Elephant National Park subpopulation. These subpopulations form distinct management units. Following the outbreaks of diseases such as bovine tuberculosis and corridor disease, South African National Parks (SANParks) recognised the need to conserve the genetic diversity of the Kruger National Park (KNP) population. To do this, a disease-free population was established in the Mokala National Park (MNP) in 1999, accordingly, an additional subpopulation is recognised, bringing the total to four. While measures of historic effective population size (Ne) have been investigated (De Jager et al. 2020), more in depth analyses is needed to calculate current Ne for these subpopulations. However, until such time, estimates can be obtained using census data.
The current census size (Nc) for MNP is approximately 400 individuals. Using the Nc:Ne (effective population size) ratio of 0.1-0.3, the Ne of this subpopulation is approximately 40-120 individuals which is below the recommended Ne of 500. In 2011 the census size for the KNP was estimated at 40,900 individuals and this would result in an Ne of 4,090-12,270 (using a ration of 0.1-0.3). The most recent census size for HiP estimated a population of 4,544 individuals. Using the same Nc: Ne ratio, the Ne has been estimated to be between 454-1,363 individuals. Lastly, a census size of 800 individuals was found for the Addo subpopulation (Zimmerman, pers. comm.) will result in an Ne of between 80-240 individuals. Most of these populations show Ne estimates which are below the recommended Ne (the KNP and HiP populations would be more resilient to future change, but the other two populations will require more conservation efforts).
Habitats and ecology
Buffalo occupy a broad range of habitats, including forest (to elevations well over 4,000 m asl), moist lowland rainforests, coastal savannahs, montane grasslands, semi-arid bushland, Acacia woodland, and Miombo Brachystegia woodland. They do not inhabit deserts and sub-deserts, for example the Namib and the Saharan/Sahelian transition zone (Prins & Sinclair 2013). Funston et al. (1994) found that the four factors that appeared to influence habitat selection of buffalo were forage availability, cover for protection against predators, proximity to water and the mobility of the herd. They drink water regularly and usually graze or take shelter in thick riverine vegetation (Skinner & Chimimba 2005). They require abundant grass, shade and water, and the absence of these requirements probably prevented their occupation of the Karoo or open grassland plains of the Highveld. Although abundant grass is required, large open floodplains or grasslands are avoided as they require the shade of trees to rest under during the hottest hours of the day. The digestive system of the buffalo is typical of bulk and roughage grazers, and is not suitable for a diet exclusively of browse material (Hofmann 1989), but they do occasionally take browse (Novellie et al. 1991; Venter & Watson 2008). They are prone to be selective of grass species and grass parts (Sinclair 1974; Hunter 1996; Prins 1996).
Buffalo are gregarious and occur in mixed herds numbering up to 3,000 individuals. In the KNP, herd numbers average 300–500 individuals (Whyte 2004) and larger herds are often associated with a higher proportion of juveniles (Tambling et al. 2013). Herds often inhabit home ranges that overlap very little with neighbouring herds and these home ranges vary in size, depending on season and the available water and forage. Old and young bulls will leave the herd and form smaller bachelor herds that tend to occupy considerably smaller ranges than the larger female-dominated herds. These smaller bachelor herds are prone to greater levels of predation as they have smaller average group sizes and tend to inhabit riskier environments (Tambling et al. 2013). Herds will normally move towards water early in the morning and again early in the evening. They are most active while feeding early in the morning and late afternoon and are characterised by considerable feeding bouts at night. Buffalo employ this activity pattern both in the presence and absence of predators; however, under high predation risk, buffalo may increase midday movement with a corresponding decrease in early morning movement (Tambling et al. 2015).
In the KNP, Pienaar (1969) recorded that calves are typically born between January and April, with a peak in January/February, coinciding with a peak in grass growth and protein content (Skinner & Chimimba 2005). Ovulation in females and spermatogenesis in males was found to commence after about 3.2 years (Sinclair 1977; Taylor 1985), and 2.5 years of age, respectively (Taylor 1985). Generally, females may give birth to their first calf when they are 4–5 years old (Carmichael et al. 1977; Taylor 1985; Mizutani 1987). Although males may reach sexual maturity between the ages of 3.5 and 5.5 years, dominant bulls typically prevent younger bulls from breeding until they are 7–8 years old (Skinner & Chimimba 2005). Over an age of 10 years, males are no longer found in breeding herds (Skinner & Chimimba 2005). A single calf is born, weighing approximately 31.1 kg for males, and 31.2 kg for females, following a mean gestation period of 340 days (Vidler et al. 1963). Calves suckle for about 9 months, and may remain with their mothers for up to two years (Skinner & Chimimba 2005).
Ecosystem and cultural services: Buffalo, as bulk grazers, are important ecosystem engineers who facilitate the presence of more selective, smaller grazers (Venter et al. 2014). They are an important food source for the larger predators like African Lion (Panthera leo) (Hayward & Kerley 2005). Buffalo and Lions (which prey on them) are both part of the “Big Five” and are important species for tourism in Africa (Okello et al. 2008). As part of the “Big Five”, they are sought after as hunting trophies. The tourism and hunting industries are of major socio-economic importance in South Africa.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Suitable | – |
| 1.6. Forest -> Forest – Subtropical/Tropical Moist Lowland | – | Suitable | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 3.4. Shrubland -> Shrubland – Temperate | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Marginal | – |
| 3.6. Shrubland -> Shrubland – Subtropical/Tropical Moist | – | Suitable | – |
| 4.5. Grassland -> Grassland – Subtropical/Tropical Dry | – | Marginal | – |
| 4.6. Grassland -> Grassland – Subtropical/Tropical Seasonally Wet/Flooded | – | Suitable | – |
| 4.7. Grassland -> Grassland – Subtropical/Tropical High Altitude | – | Suitable | – |
| 5.4. Wetlands (inland) -> Wetlands (inland) – Bogs, Marshes, Swamps, Fens, Peatlands | – | Suitable | – |
Life History
Generation Length: (Not specified)
Age at maturity: female or unspecified: Cows begin ovulating at about three years of age and have their first calf at 4-5 years of age (Prins and Sinclair 2013)
Age at Maturity: Male: 5-6 years of age, but unlikely to breed until at least 7 years of age
Size at Maturity (in cms): Female: shoulder height 145 cms, weight 446 kg
Size at Maturity (in cms): Male: shoulder height 144 weight 750 kg
Longevity: In the wild about 20 years and in captivity up to 29 years
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth: about 30 kg
Gestation Time: 340 days
Reproductive Periodicity: Birth peak in region over the summer months
Average Annual Fecundity or Litter Size: One
Natural Mortality: (Not specified)
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: A gregarious species found in stable breeding herds, bachelor herds and lone bulls.
Systems
System: Terrestrial
General Use and Trade Information
Since disease became a major threat to buffalo populations, the breeding of disease-free buffalo in South Africa has become a highly lucrative industry. Auction sales prices of buffalo have increased significantly over the years (Power 2014). Current average prices for live buffalo are between R180,000 and R450,000 per animal. There is also an increasing trend to sell animals as “stud” breeding stock when they have large, trophy quality horns. Prices of up to R44 million have recently been paid for some of these “stud” bulls. Cows are traded in a similar way and also reach high prices at game auctions in South Africa. There is considerable trophy hunting taking place of both diseased and disease-free buffalo. The current trophy hunting value is between R80,000 and R350,000 depending on the client and trophy quality. Live animals have been sold on auction from the smaller Addo Elephant National Park (Addo) for more than 10 years and, based on recent estimates, this trade is unlikely to have any impact on the Addo subpopulation (C. Tambling unpubl. data). Similarly, buffalo are auctioned from provincial reserves in the Eastern Cape and the North West Provinces (Nel 2015), as well as from current breeding operations. Overall, this trade is unlikely to affect the population in the assessment region. Buffalos are also used for bushmeat (Lindsey et al. 2012). However, the bushmeat trade, in many cases, has reached a commercial level in Africa and may not be considered subsistence-use anymore.
Wildlife ranching and the private sector have generally had a positive effect on this species as it has been widely reintroduced onto private properties within its natural distribution and introduced to those outside of its distribution. Private landowners have also bred numerous disease-free herds that can be reintroduced into protected areas. There is an increasing trend to breed buffalo under intensive conditions due to the high value and demand attached to the species. However, the selective breeding of buffalo for “stud” purposes could pose a significant threat to the population’s genetic integrity. Inbreeding and selective breeding could cause reduced genetic diversity with subsequent negative effects on the population. More species-specific research is needed to confirm or refute this. These captive subpopulations may not be suitable for re-integration with wild stock. However, there is no current need for reintroductions, as the wild stock is healthy. Reintroducing buffalo onto reserves with Lions would be counterproductive, as the few management interventions that have combined naïve buffalo with Lions have resulted in high initial levels of Lion predation. If the buffalo population is not large enough to absorb the initial predation, the reintroduced population may not persist.
The driver in the wildlife ranching industry appears to be trophy size and bulls with exceptional horns spread and large boss have fetched up to R 176 million on auction (about USD 11 million). These exceptional bulls are purchased with the aim of increasing the trophy value of his new owner’s herd. In 2024 the market price of an average mature buffalo bull for safari hunting was at least $10,000 in the Mpumalanga Lowveld. In 2024, over 1,000 live “clan’ buffalo were sold on auction is South Africa.
| Subsistence: | Rationale: | Local Commercial: | Further detail including information on economic value if available: |
| Yes | Bushmeat | Yes | Bushmeat, biltong hunting, trophy hunting, live game sales |
National Commercial Value: Yes
International Commercial Value: Yes
| End Use | Subsistence | National | International | Other (please specify) |
| 1. Food – human | true | true | – | – |
| 15. Sport hunting/specimen collecting | – | true | true | – |
Is there harvest from captive/cultivated sources of this species? Yes
Harvest Trend Comments: Trophy hunting, biltong hunting, live game sales
Threats
Historically, buffalo subpopulations suffered most severely in the 1890s, due to the great rinderpest epidemic, which was associated with pleuro-pneumonia and resulted in mortalities of up to 95% among wild ungulates and livestock (Winterbach 1998). Rinderpest, anthrax and other diseases persisted, causing localised declines and extinctions of subpopulations throughout the 20th century, as rinderpest spread from cattle to wildlife. However, the World Organisation for Animal Health declared rinderpest completely eradicated worldwide in May and June 2011 (World Organisation for Animal Health, accessed August 2014). Within the assessment region, the strict controls in regulating buffalo movement have resulted from the risk that buffalo pose to cattle and vice versa through disease transmission. Bovine Tuberculosis is of primary concern, particularly in the KNP and the Kwa-Zulu Natal Province. Tuberculosis was first diagnosed in buffalo in KNP in 1990 (Michel et al. 2006), although the growth rate and demographics of the subpopulation are unchanged (Cross et al. 2009). Over the past years, the disease has spread northwards (Michel et al. 2006). Potential negative long-term effects include the threat to the survival of threatened species that come into contact with infected buffalo, the risk of spill-over to neighbouring communal cattle, which could affect human health, and negative economic impacts caused by national and international trade restrictions (Michel et al. 2006). The population at Addo Elephant National Park is still completely disease-free (no confirmed cases of diseases of economic concern) and buffalo are sold at auctions each year. Most populations within formally protected areas outside of the veterinary red line (the hypothetical line separating diseased buffalo populations from disease-free populations) originated from this Addo population. The KNP population is also infected with foot and mouth disease, corridor disease and brucellosis, and is behind the veterinary red line. Additionally, a major population in Hluhluwe-iMfolozi Park is infected with Tuberculosis and corridor disease. Through human error, the population in Madikwe has been infected with Bovine Tuberculosis and that in Loskop dam Nature Reserve with corridor disease. Corridor disease has been regarded as self-limiting in cattle because they usually die before the parasite develops to the piroplasm stage, which is infective to ticks. Experimental evidence suggests that some bovines survive the disease, however, and may serve as reservoirs of infection. Furthermore, South Africa is considered free of T. parva, except in designated Corridor disease-infected areas including and bordering the Kruger National Park and Hluhluwe-iMfolozi Park. Therefore, the national herd is essentially naïve and completely susceptible to T. parva. The treatment of cattle to Theileriosis is prohibited because of the risk that subclinical cattle, showing no symptoms, may still be carriers and spread the disease (Yusufmia et al. 2010).
Buffalo are preferred target species for meat hunters and poachers. Snaring may affect local subpopulations within the assessment region (for example, Nel 2015). Poaching for bushmeat has become a major threat across the continent, and is one of the important causes of population declines in large ungulates in protected areas in other parts of Africa (Craigie et al. 2010; Lindsey et al. 2012), such as the Serengeti National Park in Tanzania, Comoé in Cote d’Ivoire, and Garamba in the Democratic Republic of the Congo. The increase in poaching is caused by an increase in the demand for bushmeat in both rural and urban areas, human encroachment into wildlife areas, lack of enforcement, lack of alternative livelihoods and food sources, lack of clear land rights, political instability and demand for traditional medicine (Lindsey et al. 2012). A significant increase in mineral and gas mining and exploration, with associated influx of people into uninhabited or low human density areas, are also considered to be a major factor causing increased poaching activity (Thibault & Blaney 2003).
Additionally, encroachment by humans and their domestic animals on the borders of protected areas causes edge effects and subsequently could have a negative influence on buffalo habitat.
Conservation
This species occurs widely across the assessment region, with the bulk of the population within protected areas, including the KNP, as well as Addo Elephant National Park and Great Fish River Nature Reserve. The latter populations contain the original Addo disease-free populations. Thus, the bulk of the population exists in well-managed formal protected areas. This species’ economic value has led to its reintroduction into several private properties across its natural distribution. The future status of this subspecies is closely linked to the future of protected areas, game ranches and well-managed hunting areas, since it is a frequent target of poachers. No immediate intervention is required. However, strategic translocations to sustain genetic diversity are necessary (Nel 2015).
Additionally, the success of this subspecies is tied to the control of diseases, especially where the transmission of disease from buffalo to cattle could occur. In terms of Regulation 20 published under the Animal Diseases Act, 1984 (Act No. 35 of 1984), no live cloven hoofed animals may be moved out of South Africa’s foot and mouth disease infected zones. Veterinary regulations have led to the restriction of movement of buffalo that are carrier of the foot and mouth virus beyond the red line in South Africa as this disease is easily transmitted to cattle and this will impact on the export of a range of agricultural products. This limits opportunities to establish this species outside its current range in South Africa. There are programmes in place to breed disease-free animals from behind the red line. This should be continued, but land managers should ensure that genetic diversity is retained and that there is no threat of inbreeding through the use of well-coordinated translocation policies. Solutions to eliminate Bovine Tuberculosis and corridor disease from infected populations could also be game changers for the species and the wildlife ranching industry.
The decline in capacity of provincial Conservation Agencies (Patel et al. 2023) is a concern for the control of movements of animals infected with Bovine Tuberculosis and Theileriosis.
Recommendations for land managers and practitioners:
- Develop this species as a cornerstone of the sustainable, wildlife-based rural economy.
- Provide incentives for landowners to create conservancies where the benefits of this species are shared.
- Responsible management of buffalo herds and breeding is encouraged.
- Strictly observe all veterinary regulations on the movement and introduction of live buffalo.
Research priorities:
- Methods of creating wildlife-based economies from this species through ranching, tourism and hunting.
- The effect of current stud breeding on the species’ genetic diversity.
- Further continued research on disease risks (especially non-indigenous diseases like Bovine Tuberculosis).
- Research the efficacy of a vaccine for Bovine Tuberculosis and the possible total elimination of corridor disease in managed herds.
- The scale, distribution and trends in buffalo bushmeat hunting, especially along the borders of protected areas; as well as the associated ecological, social and economic impacts.
Encouraged citizen actions:
- Landowners should create conservancies for this species and engage local stakeholders to create sustainable, wildlife-based rural economies.
- Lobby government to address lack of alternative livelihoods and food sources with innovative sustainable development solutions (e.g. encouraging smaller families, reducing reliance on natural resources, effective land-use planning, etc.).
Bibliography
Carmichael DA, Patterson L, Drager N, Breton DA. 1977. Studies on reproduction in the African buffalo (Syncerus caffer) in Botswana. South African Journal of Wildlife Research 7: 45-52.
Clinning G. 2012. Hluhluwe-iMfolozi Park Game Census Report 2012/2013. Unpublished Report. Ezemvelo KZN Wildlife, South Africa.
Craigie, I.D., Baillie, J.E.M., Balmford, A., Carbone, C., Collen, B., Green, R.E. and Hutton, J.M. 2010. Large mammal population declines in Africa’s protected areas. Biological Conservation 143: 2221-2228.
Du Toit, J.G. 2001. The African buffalo. Game ranch management (4th edition). Edited by J du P Bothma. Van Schaik Publishers, Pretoria, South Africa, pp.182-185.
Du Toit J.G. 2005. The African savanna buffalo. Intensive wildlife production in southern Africa. Edited by J du P Bothma and N van Rooyen. Van Schaik Publishers, Pretoria, South Africa, pp.78-105.East, R. (compiler). 1999. African Antelope Database 1998. IUCN, Gland, Switzerland and Cambridge, UK.
Ferreira S, Gaylard, A, Greaver, C, Hayes, J, Cowell C, Ellis G. 2013. Animal abundances in Parks 2012/2013. Scientific Services, SANParks, Skukuza, South Africa.
Funston PJ, Mills MGL. 2006. The influence of lion predation on the population dynamics of common large ungulates in the Kruger National Park. South African Journal of Wildlife Research 36: 9-22.
Funston PJ, Skinner JD, Dott HM. 1994. Seasonal variation in movement patterns, home range and habitat selection of buffaloes in a semi-arid habitat. African Journal of Ecology 32: 100-114.
Hayward MW, Kerley GI. 2005. Prey preferences of the lion (Panthera leo). Journal of Zoology 267: 309-322.
The Heinz Center. 2012. Climate-change Vulnerability and Adaptation Strategies for Africa’s Charismatic Megafauna. Washington, DC, 56 pp.
Hofmann RR. 1989. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: a comparative view of their digestive system. Oecologia 78: 443-457.
Hunter CG. 1996. Land uses on the Botswana/Zimbabwe border and their effects on buffalo. South African Journal of Wildlife Research 26: 136-150.
IUCN SSC Antelope Specialist Group. 2008. Syncerus caffer. The IUCN Red List of Threatened Species 2016: e.T21251A9260904.
Laubscher, L. & Hoffman, L. 2012. An Overview of Disease-Free Buffalo Breeding Projects with Reference to the Different Systems Used in South Africa. Sustainability 2012, 4, 3124-3140.
Le Roex N, Noyes H, Brass A, Bradley DG, Kemp SJ, Kay S, Van Helden PD, Hoal EG. 2012. Novel SNP discovery in African buffalo, Syncerus caffer, using high-throughput sequencing. PloS One 7: e48792.
Michaux, J., Smitz, N., Van Hooft, P. 2023. Taxonomic status of the African Buffalo. In: Ecology and Management of the African Buffalo. Eds. Caron, A., Cornelis, D., Chardonnet, P., Prins, HHT., Cambridge University Press, Cambridge, United Kingdom, pp. 49-65.
Mizutani F. 1987. Behaviour and patterns of maternal investment in African buffalo Syncerus caffer (Sparrman). University of Zimbabwe.
Nel, P. 2015. Population estimates for large herbivores and predators in protected areas in the North West Parks Board November 2015. North West Parks Board.
Novellie P, Hall-Martin AJ, Joubert D. 1991. The problem of maintaining large herbivores in small conservation areas: deterioration of the grassveld in the Addo Elephant National Park. Koedoe 34: 41-50.
Okello MM, Manka SG, D’Amour DE. 2008. The relative importance of large mammal species for tourism in Amboseli National Park, Kenya. Tourism Management 29: 751-760.
Pacifici, M., Santini, L., Di Marco, M., Baisero, D., Francucci, L., Grottolo Marasini, G., Visconti, P. and Rondinini, C. 2013. Generation length for mammals. Nature Conservation 5: 87–94.
Patel. T, O. Cowan, I. Little, Y. Friedmann, A. Blackmore. 2023. The State of Provincial Reserves in South Africa, Challenges and Recommendations. https://ewt.org/wp-content/uploads/2023/04/Provincial-Reserve-Management-Report-2023-FINAL-Print.pdf
Peinke DM, Gibisela Z. 2014. Game management recommendations for 2014. Eastern Cape Parks and Tourism Agency, East London.
Pienaar U de V. 1969. Observations on developmental biology, growth and some aspects of the population ecology of African buffalo (Syncerus caffer caffer Sparrman) in the Kruger National Park. Koedoe 12: 29-53.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Prins HHT, Sinclair ARE. 2013. Syncerus caffer. In: Kingdon JS, Hoffmann M (ed.), The Mammals of Africa. Volume VI: Pigs, Hippopotamuses, Chevrotain, Giraffes, Deer and Bovids., pp. 125-136. Bloomsbury Publishing, London, UK.
Prins, H.H.T. 1996. Behaviour and Ecology of the African Buffalo: Social Inequality and Decision Making. Chapman & Hall, London, UK.
Sinclair ARE. 1974. The natural regulation of buffalo populations in East Africa: the food supply as a regulating factor, and competition. African Journal of Ecology 12: 291-311.
Sinclair ARE. 1977. The African buffalo: a study of resource limitation of populations. University Press, Chicago, USA.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Tambling CJ, Ferreira SM, Adendorff J, Kerley GI. 2013. Lessons from management interventions: consequences for lion-buffalo interactions. South African Journal of Wildlife Research 43: 1-11.
Tambling CJ, Minnie L, Meyer J, Freeman EW, Santymire RM, Adendorff J, Kerley GIH. 2015. Temporal shifts in activity of prey following large predator reintroductions. Behavioral Ecology and Sociobiology 69: 1153-1161.
Taylor RD. 1985. The response of buffalo, Syncerus caffer (Sparrman), to the Kariba lakeshore grassland (Panicum repens L.) in Matusadona National Park. Ph.D. Thesis. University of Zimbabwe, Harare, Zimbabwe.
Taylor WA, Lindsey PA, Davies-Mostert HT. 2015. An assessment of the economic, social and conservation value of the wildlife ranching industry and its potential to support the green economy in South Africa. Green Economy Research Report, Endangered Wildlife Trust, Johannesburg, South Africa.
Thibault, M. Blaney, S. 2003. The Oil Industry as an Underlying Factor in the Bushmeat Crisis in Central Africa. Conservation Biology 17: 1807-1813.
Venter JA, Prins HH, Balfour DA, Slotow R. 2014. Reconstructing grazer assemblages for protected area restoration. PloS One 9: e90900.
Venter JA, Watson LH. 2008. Feeding and habitat use of buffalo (Syncerus caffer caffer) in the Nama-Karoo, South Africa. South African Journal of Wildlife Research 38: 42-51.
Venter JA. 2006. The feeding ecology of Buffalo (Syncerus caffer caffer) in Doornkloof Nature Reserve, Northern Cape Provinve, South Africa. M.Sc. Thesis. Nelson Mandela Metropolitan University, Port Elizabeth, South Africa.
Vidler BO, Harthoorn AM, Brocklesby DW, Robertshaw D. 1963. The gestation and parturition of the African buffalo (Syncerus caffer caffer Sparrman). African Journal of Ecology 1: 122-123.
Whyte IJ. 2004. Census results for elephant and buffalo in the Kruger National Park between 1997 and 2004. Scientific Report 03/04 Internal Report. Kruger National Park, South African National Parks, Pretoria, South Africa.
Winterbach, H. E. K. 1998. Research review: the status and distribution of Cape buffalo Syncerus caffer caffer in southern Africa. South African Journal of Wildlife Research 28(3): 82-88.
de Vos V, Bengis RG, Kriek NPJ, Michel A, Keet DF, Raath JP, Huchzermeyer HFKA. 2001. The epidemiology of tuberculosis in free-ranging African buffalo (Syncerus caffer) in the Kruger National Park, South Africa. The Onderstepoort Journal of Veterinary Research 68: 119.
van Rooyen N. 2004. Vegetation types and wildlife re-establishment in the Greater St Lucia Wetland Park. Unpublished Report. Isimangaliso Wetland Park, St Lucia, South Africa.
Yusufmia. S.B.F.S ; N E Collins; R Nkuna; M Troskie; P Van den Bossche; B L Penzhorna. 2010. Occurrence of Theileria parva and other haemoprotozoa in cattle at the edge of Hluhluwe-iMfolozi Park, KwaZulu-Natal, South Africa.