

White-tailed Mongoose
Ichneumia albicauda

2025 Red list status
Least Concern
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Ichneumia albicauda – (G. Cuvier, 1829)
ANIMALIA – CHORDATA – MAMMALIA – CARNIVORA – HERPESTIDAE – Ichneumia – albicauda
Common Names: White-tailed Mongoose (English), Witstertmuishond (Afrikaans), Ubuchakide (Ndebele), Jerenyenje (Shona), Liduha (Swazi), Tlolota (Tsonga), Lesalamotlhaka, Lesêlamotlhaka, Mokala, Mosalamotlhaka, Mosêlamotlhaka, Sesêlamotlhaka (Tswana), Mutsherere (Venda), Ingqwalashu (Xhosa), Gqalashu (Zulu)
Synonyms: Herpestes albicauda G. Cuvier, 1829
Taxonomic Note:
This species is occasionally mistaken with Meller’s Mongoose (Rhynchogale melleri), which also sometimes has a white tail. However, the White-tailed Mongoose is usually larger, and its body appears black, rather than brown (Skinner and Chimimba 2005). Further confusion in identification is sometimes created by the fact that Ichneumia albicauda individuals with black tails have been recorded in several areas of the African distribution range (Pringle 1977; Dehgani et al. 2008; Do Linh San et al. 2016). This colour polymorphism observed in some Western populations in African populations reflects variation at the individual level (Dehgani et al. 2008).
Only one subspecies has been listed, from southern Africa, I. a. grandis (Thomas 1890; Dehgani et al. 2008), but the nominate form has a wide distribution across much of the rest of Africa (Meester et al. 1986). There are believed to be three large genetic clades which may be congruent with some of the recognised subspecies across Africa (Dehgani et al. 2008). The White-tailed Mongoose probably originated in southern Africa, from where it dispersed northwards and colonized eastern and western parts of Africa, as well as the Arabian Peninsula (Dehgani et al. 2008).
Molecular systematics and morphological studies have confirmed that this true mongoose is in the family, Herpestidae (Veron et al. 2022).
Red List Status: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Streicher, J.P.1,2,3, Power, R.J.4 & da Silva, J.M.1
Reviewers: Nicholson, S.K.5 & Do Linh San, E.6
Institutions:1South African National Biodiversity Institute,2BirdLife South Africa,3University of KwaZulu-Natal,4North West Provincial Government,5Endangered Wildlife Trust, 6Sol Plaatjie University
Previous Assessors and Reviewers: Do Linh San, E., Stuart, C. & Stuart, M.
Previous Contributors: Madikiza, Z. & Relton, C.
Assessment Rationale
The White-tailed Mongoose is listed as Least Concern as it is common across the assessment region and exists in a wide range of habitat types – even in human-modified habitats where they feed opportunistically. Therefore, there is little reason to believe that they are threatened.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Streicher JP, Power RJ & da Silva JM. 2025. A conservation assessment of Ichneumia albicauda. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The White-tailed Mongoose is widespread in sub-Saharan Africa from Senegal and Gambia to the Horn of Africa, and then southwards to southeast South Africa (Taylor 2013). Extralimital to the African continent, it is present along much of the coast of Saudi Arabia to Oman and also recorded from Farasan Kabir Island in the Red Sea, where they may have been introduced (Taylor 2013). It ranges from sea level to 3,500 m asl in Ethiopia (Admasu et al. 2004).
In southern Africa, it ranges marginally into the extreme northeastern parts of Namibia, in northern Botswana, and through most of Zimbabwe and Mozambique, aside from the more arid regions (Skinner & Chimimba 2005). Within the assessment region, the species still occurs in the eastern lowveld of Limpopo and Mpumalanga, southwards to KwaZulu-Natal, the Free State and the Eastern Cape as far as Addo Elephant National Park. The earlier occurrence baselines, prior to the 1990s, were KwaZulu-Natal (Pringle 1977; Rowe-Rowe 1978), Eastern Cape (Stuart 1981), south-eastern Northern Cape (Stuart 1981), Free State (Lynch 1983), and northern provinces of Mpumalanga and Limpopo (Rautenbach 1982), with increasingly more evidence from the North West into the 2000s (Power et al. 2019) In the Verloren Vallei Nature Reserve, between 2015 and 2019, White-tailed Mongoose were not detected by camera traps (Loock & Swanepoel 2019), though the species appears to be regionally present (Figure 1).
In the North West Province, the species is thought to have significantly expanded its range westward into the Kalahari (Power 2014), and has been recorded progressively further east with occurrences reported in 2016 in the !Khamab Kalahari Reserve (Mann et al. 2016). Given the amount of survey effort expended in the vegetation type of the area (see Power et al. 2019), it is likely that this record could be a range expansion westward (see Figure 1).
Their presence in Eswatini is currently unknown.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): 3,500m m asl
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
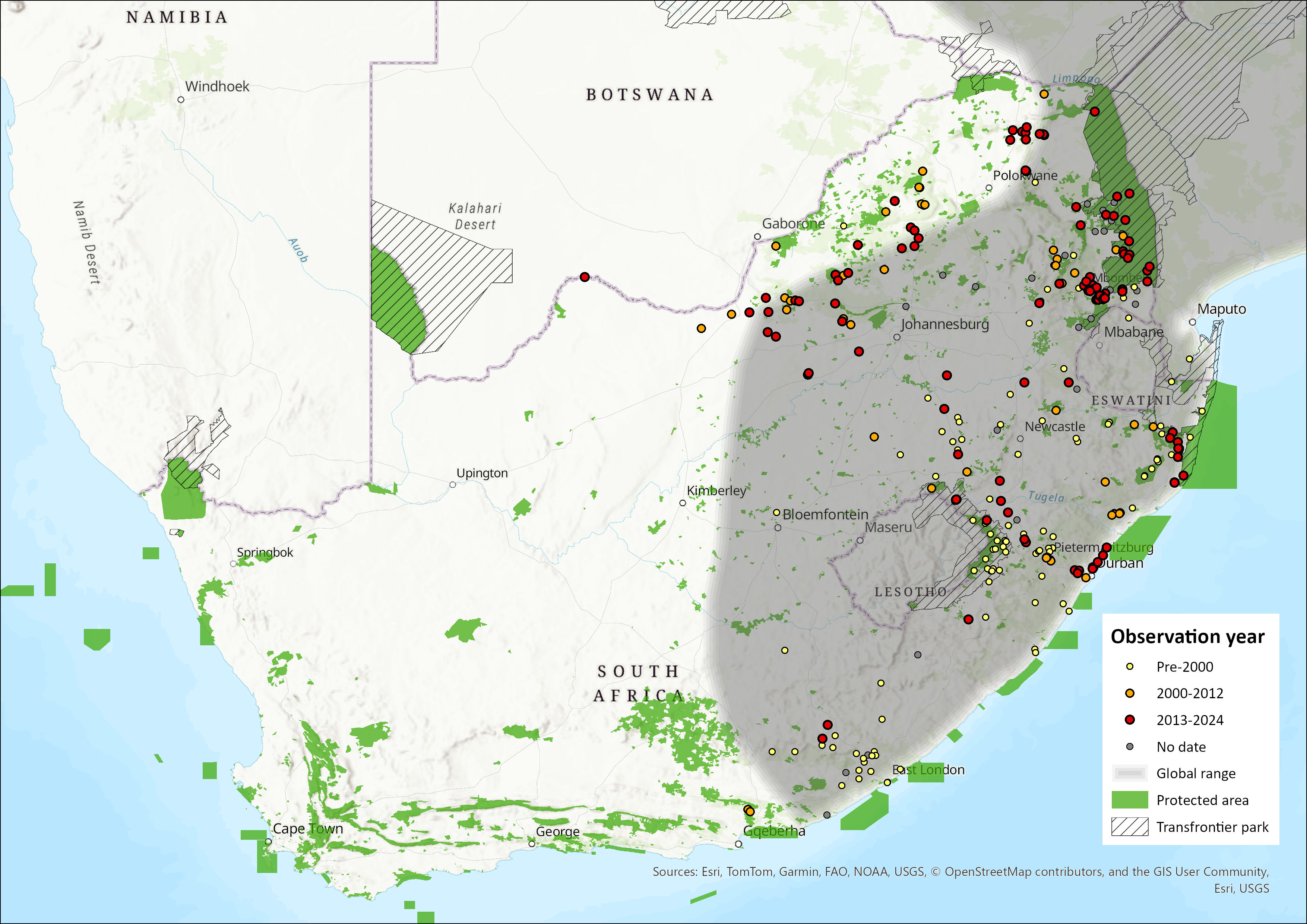
Figure 1. Distribution records for White-tailed Mongoose (Ichneumia albicauda) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical, Palearctic
Occurrence
Countries of Occurrence
Surprisingly, in the mid-1990s, White-tailed Mongoose had not been recorded in Eswatini on spotlight surveys (Monadjem et al. 1998). It is unknown whether they have subsequently been recorded.
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Benin | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Burkina Faso | Extant | Native | – | – |
| Cameroon | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Chad | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Côte d’Ivoire | Extant | Native | – | – |
| Djibouti | Extant | Native | – | – |
| Egypt | Extant | Native | – | – |
| Eritrea | Extant | Native | – | – |
| Eswatini | Unknown | – | – | – |
| Ethiopia | Extant | Native | – | – |
| Gambia | Extant | Native | – | – |
| Ghana | Extant | Native | – | – |
| Guinea | Extant | Native | – | – |
| Guinea-Bissau | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Lesotho | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mali | Extant | Native | – | – |
| Mauritania | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Niger | Extant | Native | – | – |
| Nigeria | Extant | Native | – | – |
| Oman | Extant | Native | – | – |
| Saudi Arabia | Extant | Native | – | – |
| Senegal | Extant | Native | – | – |
| Sierra Leone | Extant | Native | – | – |
| Somalia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| South Sudan | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Yemen | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Given the advent of climate change may see increased woody plant cover (Foden and Young 2016) and given their predilection for well bushed habitat (Ehlers-Smith et al. 2017), this may favour White-tailed Mongoose expansion.
Population Information
This species is considered common, and in some parts of its southern African range is considered one of the most common small carnivores in some landscapes (Skinner & Chimimba 2005). Densities as high as 4.3 individuals / km² were reported from the Serengeti (Waser 1980), however this may not reflect the South African situation.
In the Madikwe Game Reserve in the North West, in 2019, White-tailed Mongoose were recorded, but with a relatively low number of detections (Smith et al. 2023). In the Associated Private Nature Reserve in the Lowveld of Limpopo, in 2019, White-tailed Mongoose were recorded, with a moderate number of detections (Smith et al. 2023). In 2017, they occurred in the Phinda area of northern KwaZulu-Natal (Da Rosa 2019). In Maputaland, when comparing the species across a protected area (Phinda), mixed game/livestock farms and a pastoral community area, the species occurred at 55%, 46% and 11% of these sites respectively (Da Rosa 2019). These suggest lowest occurrence in a pastoral community area. Since the species is not individually recognisable, one may rely on relative abundance indices for the species across its range (Da Rosa 2019; Smith et al. 2023).
Considering the relatively wide distribution of this species and a minimum average density of at least 0.1 individual / km², we estimate that there are well over 10,000 mature individuals in the assessment region (Do Linh San et al. 2016). The small female home-ranges estimated in KwaZulu-Natal (i.e. 0.7 km²) (Streicher et al. 2020), suggests high densities can be attained in suitable habitats, even those relatively modified by humans (see Streicher et al. 2020).
Population Information
Current population trend: Unknown, but probably stable based on wide extent of occurrence and lack of threats.
Continuing decline in mature individuals: Unknown, but probably not.
Number of mature individuals in population: Probably > 10,000.
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: It is not currently possible to determine the extent or number of subpopulations.
Severely fragmented: No. This species can exist in multiple habitats, including agricultural lands, rural gardens and urban areas. Additionally, its range may be increasing with the rise of wildlife ranching.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
No population genetic studies have been conducted on the White-tailed Mongoose; however, a phylogenetic study based on partial sequences of the mitochondrial control region was able to identify three geographically associated genetics clusters: a southern clade (South Africa); a western clade (Guinea); and an eastern clade (United Arab Emirates, Yemen, Saudi Arabia, Ethiopia, Central African Republic, Kenya and Uganda) (Dehgani et al. 2008). These clades lend support to some of subspecies identified based on morphological evidence (Taylor, 1972; Coetzee, 1977). Based on this evidence and considering the species is common across some landscapes, it is assumed that a single Evolutionary Significant Unit (ESU) or metapopulation exists within the assessment region. However, further genetic investigations are needed, with increased sampling, to uncover whether additional genetic structure exists and to estimate effective population sizes.
While accurate estimates of Ne are not available, it can be inferred based on the assumption that over 10,000 individuals exist within the assessment region. Applying a Ne/Nc conversion ratio between 0.1-0.3, Ne is estimated between 1,000-3,000 individuals, far exceeding the 500 threshold for a stable and healthy population.
Habitats and ecology
The White-tailed Mongoose occurs in well-watered grasslands, savannah and woodland areas, but is absent from moist forested areas, high altitudes above the treeline, deserts and semi-deserts (Taylor 2013). Generally preferring humid or sub-humid areas, this species is restricted from the drier expanses of the Succulent and Nama Karoo biomes of the assessment region (Skinner & Chimimba 2005). They are also found in grassland or thornveld, and sometimes in association with permanent water, such as streams (Stuart 1981). However, they have been found in relatively arid areas, in particular the Eastern Kalahari Bushveld (after Mucina & Rutherford 2006) in the North West province too.
It is adaptable to some human-modified habitats, with individuals often found around towns and villages where they feed opportunistically on garbage (Taylor 2013), and associated croplands in agricultural areas in Limpopo (Williams et al. 2018), and KwaZulu-Natal (Ramesh and Downs 2015; Streicher et al. 2020). In Oman it has been found associated with gardens and plantations (Harrison & Bates 1991). The species does prefer bush over and above forest (Ehlers-Smith et al. 2017). Some studies have suggested they are deterred by livestock areas (Ramesh & Downs 2015), while others have suggested they are strongly associated with livestock ranging areas (Wiliams et al. 2018), likely related to an association with invertebrate food and livestock dung (see Skinner & Chimimba 2005; Williams et al. 2018). Large predators do kill White-tailed Mongoose (Caro and Stoner 2003), so intra-guild competion and predation readily occurs, and may impact their behavioural ecology. On gme and livestock farms in Maputaland, White-tailed Mongoose abundance was related to rodent abundance, tree cover and distant from cattle presence (Da Rosa 2019, but see Williams et al. 2018).
This species is primarily insectivorous. Termites, grasshoppers, crickets, beetles and beetle larvae constitute a high percentage of its diet (Rowe-Rowe 1978; Smithers & Wilson 1979; Stuart 1981; Rautenbach 1982). Amphibians, reptiles and murid rodents may also supplement its diet (Skinner & Chimimba 2005). Although the White-tailed Mongoose is nocturnal, with active behaviour extending well into the night, occasional diurnal behaviour is not unheard of (Stuart 1981). In Ethiopia, resting sites during the day were located amongst rocks or even in empty buildings (Admasu et al. 2004), but termitaria, disused Aardvark (Orycteropus afer) or Springhare (Pedetes capensis) holes are also commonly utilised as dens (Skinner & Chimimba 2005).
This species is primarily solitary, but small family groups or pairs are occasionally observed at night (Skinner & Chimimba 2005). In an agricultural landscape of the Ethiopian Highlands, a study into the spatial ecology of this species found the average size of three adult male home ranges to be 3.2 km², while an individual female home range was recorded as 2.6 km² (Admasu et al. 2004). The male home ranges showed very little (2%) overlap with one another, however, the female’s home range was found to overlap with those of the two adjacent male home ranges by 81% and 25%, respectively (Admasu et al. 2004). A home range in Kenya was recorded at 8 km² (Baker 1997). Similarly, a study in the KwaZulu-Natal Midlands, found the average size of two female home ranges to be 0.7 ± 0.10 km² (Streicher et al. 2020). Little is known of the reproductive seasonality of this species. However, in Zimbabwe pregnant and lactating females were recorded from October to February (Shortridge 1934); and Rowe-Rowe (1978) recorded both a lactating female and a pregnant female in November in KwaZulu-Natal. Litter size is suggested to be 1–4 young (Taylor 2013), but Skinner and Chimimba (2005) mentioned an average litter size of 1.4. The gestation period is estimated at 2 months (Baker 1997).
Ecosystem and cultural services: This species may predate on pest species, for example, a captive female White-tailed Mongoose was known to predate on House Rats (Rattus rattus) up to 180 g in mass (Smithers 1983). Rodent remains were found in 18% of scats/stomachs of white-tailed mongoose in a pastoral agro-ecosystem of north-eastern Limpopo (Williams et al. 2018), suggesting importance as a rodentivore. However, of community respondents, 3.4-15.9% knew White-tailed Mongoose fed on rodents (Williams et al. 2018). However, up to 77.5% still perceived them to impact negatively, which suggests an underappreciation of their value in this agro-ecosystem (Williams et al. 2018).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 4.5. Grassland -> Grassland – Subtropical/Tropical Dry | – | Suitable | – |
| 14.3. Artificial/Terrestrial -> Artificial/Terrestrial – Plantations | – | Marginal | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Marginal | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Period: Estimated at 2 months (Baker 1997)
Reproductive Periodicity: (Not specified)
Litter Size: 1–4 young (Taylor 2013), but Skinner and Chimimba (2005) mentioned an average litter size of 1.4.
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
Although this species is not known to be utilised as bushmeat within the assessment region, Taylor (1972) suggested that humans were probably an opportunistic predator of adult White-tailed Mongooses. There may be limited or opportunistic use of White-tailed Mongoose skins for traditional ceremonial purposes in Venda villages of the Limpopo Province (C. Stuart & M. Stuart pers. obs. 1985-1986). In southern Benin, White-Tailed Mongoose are present in markets suggests they are used in traditional medicine (Djagoun et al. 2023), though they are not prominent in South Africa. However, at the Faraday Market in Gauteng in 2005, traders interviewed were stocking body parts of this species for sale (Whiting, Williams & Hibbits 2016).
The species does not appear in the Professional Hunters register and there seems to be no real demand as a trophy animal in the hunting industry.
Not unlike other rarely seen nocturnal mammals, there is interest in observing them as night by select ecotourists.
| Subsistence: | Rationale: | Local Commercial: | Further detail including information on economic value if available: |
| Yes | – | – | – |
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
| End Use | Subsistence | National | International | Other (please specify) |
| 3. Medicine – human & veterinary | – | – | – | – |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
There are no major threats to the species, although they may be caught incidentally in predator control programmes principally aimed at Black-backed Jackal (Lupulella mesomelas) and Caracal (Caracal caracal) (Taylor 2013), as is by-catch in snares for bushmeat also.
In the North West Province there have been 6 White-tailed Mongoose road kills recorded since 2019, in the last 5 years (North West Directorate of Biodiversity Management records, 2019- 2023). Between April 2022 and March 2024, a driven systematic roadkill circuit of 150 km connecting the towns of Lichtenburg, Ottoshoop and Mafikeng recorded only one White-tailed Mongoose roadkill and this circuit was driven on 76 days (North West Biodiversity Management, Unpublished records). The low rate of road kills may be a function of abundance or behaviour on the part of these animals. Despite this, roadkill mortality may be an important emerging threat impacting their populations, particularly in human-impacted environments.
Domestic dogs are a possible threat, as they are indirectly negatively impacted by them (see Williams et al. 2018). Habitat loss, or degradation, may be threat, though agricultural transformation may be limited as they still readily utilise these landscapes (Ramesh & Downs 2015; Williams et al. 2018).
Conservation
The White-tailed Mongoose is present in many protected areas across its range, and it has benefitted from the prevalence of protected areas in the north-east of South Africa (Siegfried and Brown 1992), associated with its known distribution range (Skinner and Chimimba 2005; Do Linh San et al. 2016). The species benefits from a network of reserves which protect the specie’s habitat (Ramesh et al. 2016).
No specific conservation interventions are currently deemed necessary within the assessment region, but monitoring is recommended. This species is likely to benefit from the expansion of protected areas to connect suitable habitat patches.
Given the undervalued role of the White-tailed Mongoose as a rodent predator in at least Limpopo, education and awareness would be encouraged for this species (see Williams et al. 2018).
Recommendations for land managers and practitioners:
- Create conservancies to protect and connect habitat.
Research priorities:
- Monitoring subpopulations to determine population size and trends.
- General studies on the biology and ecology of this species in different habitat types.
- Assess whether range expansion has indeed occurred into the arid west
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas. As confusion with other mongoose species is possible, a photograph is required for confirmation of identification.
Bibliography
Admasu E, Thirgood SJ, Bekele A, Laurenson MK. 2004. Spatial ecology of white-tailed mongoose in farmland in the Ethiopian highlands. African Journal of Ecology 42:153–159.
Baker C. 1997. White-tailed Mongoose Ichneumia albicauda. Page 215 in Mills G, Hes L. The Complete Book of Southern African Mammals. Struik, Cape Town, South Africa.
Caro TM, Stoner CJ. 2003. The potential for interspecific competition among African carnivores. Biological Conservation, 110(1): 67-75.
Da Rosa BRP. 2019. Optimizing Small Mammal Relative Abundance Measures Using Non-Invasive Sampling and Assessment of its Contribution to Occupancy Modelling of Small Carnivores in Dry Woodland Savannah of South Africa. Master’s thesis, University of Lisbon, Lisbon.
Dehghani R, Wanntorp L, Pagani P, Källersjö M, Werdelin L, Veron G. 2008. Phylogeography of the white‐tailed mongoose (Herpestidae, Carnivora, Mammalia) based on partial sequences of the mtDNA control region. Journal of Zoology 276(4): 385-393.
Djagoun CAMS, Zanvo S, Azihou F, Nago G, Djagoun J, Vodouhê F, et al. 2023. Assessing the impact of the wildlife trade in West Africa (Benin): Functional diversity matters too. Global Ecology and Conservation 47:e02630.
Do Linh San E, Stuart C, Stuart M. 2016. A conservation assessment of Ichneumia albicauda. In Child MF, Roxburgh L, Do Linh San E, Raimondo D, Davies-Mostert HT, editors. The Red List of Mammals of South Africa. Swaziland and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Ehlers-Smith YC, Ehlers-Smith DA, Ramesh T, Downs CT. 2017. The importance of microhabitat structure in maintaining forest mammal diversity in a mixed land-use mosaic. Biodiversity and Conservation 26: 2361-2382.
Foden, W.B. and Young, B.E. (eds.) 2016. IUCN SSC Guidelines for Assessing Species’ Vulnerability to Climate Change. Version 1.0. Occasional Paper of the IUCN Species Survival Commission No. 59. Cambridge, UK and Gland, Switzerland: IUCN Species Survival Commission. x+114pp.
Harrison DL, Bates PJJ. 1991. The Mammals of Arabia. Harrison Zoological Museum, Sevenoaks, UK.
Loock D, SwanepoeL L. 2019. Assessing local patterns in carnivore occupancy and richness by camera trapping: a monitoring of diversity at Verloren Vallei Nature Reserve, Dullstroom, Mpumalanga. Wildlife & Resource Association (WRA), Bloemfontein.
Lynch CD. 1983. Mammals of the Orange Free State. Memoirs van die Nasionale Museum, Bloemfontein 18: 1-218.
Mann G, Thomas L, Kilian H, Balme G. 2016. South African Leopard Monitoring Project: Khamab Kalahari Reserve Camera-trap Survey 2016. Internal Report, Panthera & SANBI, Cape Town.
Meester JA, Rautenbach IL, Dippenaar NJ, Baker CM. 1986. Classification of southern African mammals. Transvaal Museum Monographs 5:1–359.
Monadjem A, Monadjem L, Putnam J. 1998. Notes and Records: Sighting frequencies of nocturnal mammals in Swaziland. African Journal of Ecology 36(3): 280-285.
Mucina I, Rutherford MC. 2006. The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19: South African National Biodiversity Institute, Pretoria.
Power RJ. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng, South Africa.
Power RJ, Van Straaten A, Schaller R, Mooke M, Boshoff T, Nel HP. 2019. An inventory of mammals of the North West Province, South Africa. Annals of the Ditsong Museum of Natural History 8: 6-29.
Pringle JA. 1977. The distribution of mammals in Natal. Part 2. Carnivora. Annals of the Natal Museum, 23(1): 93-115.
Ramesh T, Downs CT. 2015. Impact of land use on occupancy and abundance of terrestrial mammals in the Drakensberg Midlands, South Africa. Journal for Nature Conservation, 23: 9-18.
Ramesh T, Kalle R, Rosenlund H, Downs CT. 2016. Native habitat and protected area size matters: Preserving mammalian assemblages in the Maputaland Conservation Unit of South Africa. Forest Ecology and Management 360: 20-29.
Rautenbach IL. 1982. The mammals of Transvaal. Ecoplan Monograph 1:1–211.
Rowe-Rowe DT. 1978. The small carnivores of Natal. Lammergeyer 25:1–48.
Shortridge GC. 1934. The Mammals of South West Africa. Volume I & II. Heinemann, London, UK.
Siegfried WR, Brown C.A. 1992. The distribution and protection of mammals endemic to southern Africa. South African Journal of Wildlife Research 22(1): 11-16.
Skinner JD, Chimimba CT. 2005. The Mammals of the Southern African Subregion. Third edition. Cambridge University Press, Cambridge, UK.
Smith K, Venter JA, Peel M, Keith M, Somers MJ. 2023. Temporal partitioning and the potential for avoidance behaviour within South African carnivore communities. Ecology and Evolution. 13(8):e10380.
Smithers RHN. 1983. The Mammals of the Southern African Subregion. University of Pretoria, Pretoria, South Africa.
Smithers RHN, Wilson VJ. 1979. Check list and atlas of the mammals of Zimbabwe Rhodesia. Trustees of the National Museums and Monuments 9:1–147.
Streicher JP, Ramesh T, Downs CT. 2020. Home range and core area utilisation of three co-existing mongoose species: large grey, water and white-tailed in the fragmented landscape of the KwaZulu-Natal Midlands, South Africa. Mammalian Biology 100(3):273–283.
Stuart CT. 1981. Notes on the mammalian carnivores of the Cape Province, South Africa. Bontebok 1:1–58.
Taylor ME. 1972. Ichneumia albicauda. Mammalian Species 12:1–4.
Taylor ME. 2013. Ichneumia albicauda White-tailed Mongoose. Pages 342–346 in Kingdon J, Hoffmann M, editors. The Mammals of Africa. Volume V: Carnivores, Pangolins, Equids and Rhinoceroses. Bloomsbury Publishing, London, UK.
Veron G, Patou ML, Jennings AP. 2022. Systematics and evolution of the mongooses (Herpestidae, Carnivora). Small Carnivores: Evolution, Ecology, Behaviour, and Conservation: 61-78.
Waser PM. 1980. Small nocturnal carnivores: ecological studies in the Serengeti. African Journal of Ecology 18:167–185.
Whiting MJ. Williams VL, Hibbits TJ. 2016. A picture of health? Animal use and the Faraday traditional medicine market, South Africa. Journal of Ethnopharmacology. 179:265-73.
Williams ST, Maree N, Taylor P, Belmain SR, Keith M, Swanepoel LH. 2018. Predation by small mammalian carnivores in rural agro-ecosystems: an undervalued ecosystem service?. Ecosystem Services 30: 362-371.