

Welwitsch’s Hairy Bat
Myotis welwitschii

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Myotis welwitschii – (Gray, 1866)
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – VESPERTILIONIDAE – Myotis – welwitschii
Common Names: Welwitsch’s Hairy Bat, Welwitsch’s Myotis, Welwitch’s Bat, Welwitsch’s Mouse-eared Bat (English), Welwitsch se langhaarvlermuis, Welwitsch-langhaarvlermuis (Afrikaans), Kafukufuku (Tshokwe, Angola)
Synonyms: Scotophilus welwetschii Roberts, 1951; venustus (Matschie, 1899); welwitschi Hayman, 1963 [incorrect subsequent spelling] (see Mammal Diversity Database 2025)
Taxonomic Note:
This species is monotypic (Meester et al. 1986; Simmons 2005). No subspecies are recognised.
Red List Status: LC – Least Concern (IUCN version 3.1)
Assessment Information
Assessors: Richards, L.R.1, Howard, A.2 & da Silva, J.M.3
Reviewer: Moir, M.3
Institutions: 1Durban Natural Science Museum, 2University of the Free State, 3South African National Biodiversity Institute, 4Stellenbosch University
Previous Assessors and Reviewers: Monadjem, A., Jacobs, D., Cohen, L., MacEwan, K., Richards, L.R., Schoeman, C., Sethusa, T. & Taylor, P.
Previous Contributors: Nicholson, S.K., Raimondo, D. & Child, M.F.
Assessment Rationale
Listed as Least Concern in view of its wide distribution (estimated extent of occurrence for the assessment region is 365,568 km2), its occurrence in multiple protected areas (including Great Limpopo Transfrontier Park), and because there are no major identified threats that could cause widespread decline. It is unknown whether it tolerates human disturbed habitats. There is no evidence to suggest it qualifies for a threatened status under criterion B because it does not appear to be limited by roost availability. However, it is an uncommon species and further field surveys, and research are necessary to delimit population size and trend, distribution and habitat selection more accurately. Once such data are available, this species should be reassessed.
Regional population effects: The range of this species is continuous across the borders of the assessment region through its occurrence in the Great Limpopo Transfrontier Park. Additionally, it has intermediate wing loading (Schoeman & Jacobs 2003), so dispersal effects are probably possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Richards LR, Howard A & da Silva JM. 2025. A conservation assessment of Myotis welwitschii. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species has been widely, but patchily, recorded over much of East Africa and southern Africa, parts of southern Central Africa, and a single West African locality in southeastern Guinea (Monadjem et al. 2020; ACR 2024). In East and southern Africa, it is sparsely distributed in the eastern regions, with scattered records distributed from Ethiopia and South Sudan in the north, through Kenya, Tanzania and Malawi, to Zambia, Mozambique, Zimbabwe and northeastern South Africa (Fahr & Ebigbo 2003; Monadjem et al. 2020; ACR 2024). It has not been recorded from Namibia or Botswana (ACR 2024). Within the assessment region, it occurs in South Africa, namely in provinces of Limpopo, Mpumalanga, Gauteng, KwaZulu-Natal and marginally in the Free State. This species has been recently recorded in Eswatini but is absent from Lesotho (Monadjem et al. 2021, Skinner & Chimimba 2005). Its current estimated extent of occurrence for the assessment region is 365,568 km2.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 49 m asl (Monadjem et al. 2024)
Elevation Upper Limit (in metres above sea level): 2,380 m asl (Monadjem et al. 2024)
Depth Lower Limit (in metres below sea level): N/A
Depth Upper Limit (in metres below sea level): N/A
Depth Zone: N/A
Map
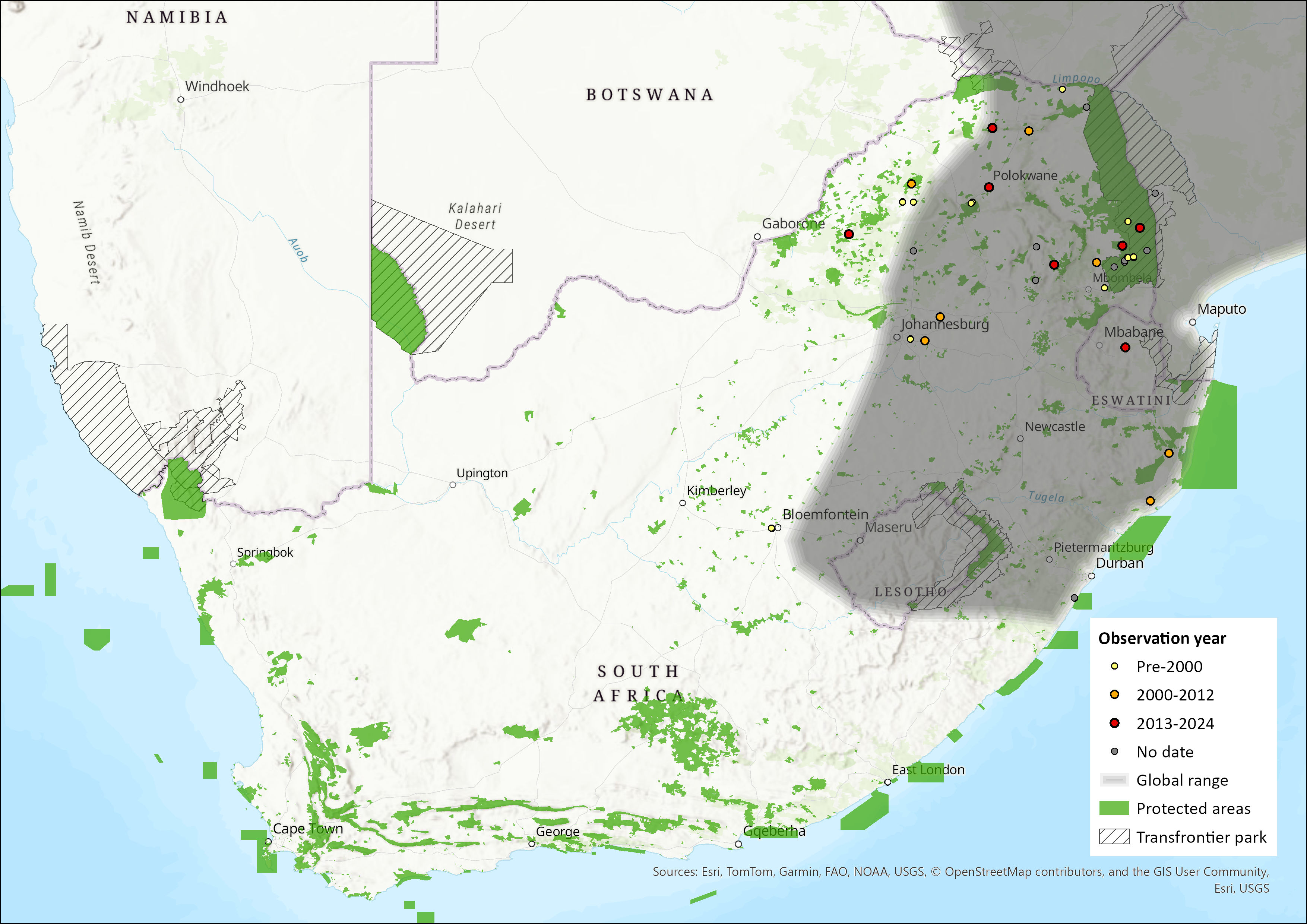 Figure 1. Distribution records for Welwitsch’s Hairy Bat (Myotis welwitschii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Welwitsch’s Hairy Bat (Myotis welwitschii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Burundi | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Eswatini | Possibly Extant | Native | – | Resident |
| Ethiopia | Extant | Native | – | – |
| Guinea | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Rwanda | Extant | Native | – | – |
| South Africa | Extant | Native | – | Resident |
| South Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: N/A
FAO Area Occurrence
FAO Marine Areas: N/A
Climate change
No formal study on the direct effects of climate change on this species has been conducted. Most studies on the effects of climate change on bats are based on predictive species distribution modelling, thus there is a lack of empirical studies measuring behavioural, physiological, phenological or genetic responses to extreme and seasonal climatic changes, especially in the Global South (Festa et al. 2023; Pio et al. 2014). Globally, there have been documented declines in bat populations, species richness and distributions in relation to water availability, with increasing global aridity, which may become a growing concern as heat waves and maximum temperatures are expected to increase over much of South Africa (Adams & Hayes 2021; Mbokodo et al. 2020).
Population Information
Although this species is widespread, it is not common (ACR 2024). It tends to roost singly, in trees and bushes (Happold 2013), and hence it is very difficult to estimate population sizes. It is poorly represented in museums, with just 30 records examined in Monadjem et al. (2020). We presume the population is stable given new – occurrence records being recorded since the previous assessment (e.g. Monadjem et al. 2021; Taylor et al. 2020; Weier et al. 2021). However, further monitoring and research is needed to estimate population size and trend.
Current population trend: Stable
Continuing decline in mature individuals? Cannot be determined
Extreme fluctuations in the number of subpopulations: Cannot be determined
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Number of mature individuals in largest subpopulation: (Not specified)
Number of Subpopulations: (Not specified)
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
A phylogenetic study on Afrotropical species of the genus Myotis identified two evolutionarily distinct lineages within M. welwitschii (Patterson et al. 2019); however, given that only a single mitochondrial gene was used and limited samples incorporated, further research employing an integrative taxonomic approach is highly recommended to better understand the evolutionary relationships within this species.
Moreover, because insufficient data exists to quantify population size, it is also highly recommended that a population genetic study be conducted to better understand genetic structure and diversity within the assessment region.
Habitats and ecology
This species mainly inhabits open woodland and savannah (Ratcliffe 2002) but has been recorded from a range of habitats including tropical dry forest, montane tropical moist forest, both dry and moist savannah, shrublands, and high-altitude grassland (ACR 2024). It may be associated with mountains covered by a woodland-forest mosaic vegetation in some areas (Fahr & Ebigbo 2003; Monadjem et al. 2020). Anecdotal observations of day roosts include factories, houses, low bushes, trees (externally in hollows and among leaves, including rolled banana leaves), and deep in caves (such as Makapan’s Cave in Limpopo) (summarised in Ratcliffe 2002). It is generally a solitary rooster (Ratcliffe 2002) and clings to surfaces in trees or shrubs (Monadjem et al. 2020). It is a clutter-edge forager and, based on limited observations in Mpumalanga, its diet comprises aerial prey such as Coleoptera and Hemiptera (Monadjem et al. 2020). Morales et al. (2024) assigned M. welwitschii to the “gleaner” ecomorph type.
Ecosystem and cultural services: This insectivorous species has been recorded from macadamia orchards (Weier et al. 2021). It is possible that it may assist with insect pest control. Worldwide, insectivorous bats contribute to controlling insect populations that damage crops (Boyles et al. 2011; Kunz et al. 2011). Ensuring a healthy population of insectivorous bats can thus decrease the need for pesticides.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Suitable | – |
| 1.9. Forest -> Forest – Subtropical/Tropical Moist Montane | – | Suitable | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 3.6. Shrubland -> Shrubland – Subtropical/Tropical Moist | – | Suitable | – |
| 3.7. Shrubland -> Shrubland – Subtropical/Tropical High Altitude | – | Suitable | – |
| 4.7. Grassland -> Grassland – Subtropical/Tropical High Altitude | – | Suitable | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Unknown | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Both sexes: Mean forearm length = 5.60 ± 0.22 cm; Mean total length = 11.60 ± 0.74 cm (Monadjem et al. 2020)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): Total body length 10,5–12,7 cm
Size at Birth (in cms): Unknown
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Unknown
Congregatory: No, records reported thus far all relate to singletons roosting in trees and bushes (Happold 2013).
Systems
System: Terrestrial
General Use and Trade Information
There is no current evidence to suggest that this species is traded or utilised however online e-commerce platforms should be monitored for illegal trade in bat taxidermy and specimens of this species (Coleman et al. 2024; Hunter et al. 2024).
Local Livelihood: None reported
National Commercial Value: Possibly contributes to insect pest control, and this has agroeconomic value (see above)
International Commercial Value: Possibly contributes to insect pest control and this has agroeconomic value (see above)
End Use: None reported
Is there harvest from captive/cultivated sources of this species? N/A
Harvest Trend Comments: N/A
Threats
There appears to be no major threats to this species (ACR 2024). It is able to utilise semi-disturbed vegetation or landscapes for roosting and foraging. However, there is ongoing habitat loss from agricultural and human settlement transformation, especially in KwaZulu-Natal (Jewitt et al. 2015) and Gauteng, which is an increasingly ongoing threat (Driver et al. 2012; GeoTerraImage 2015). Selective logging of trees for fuelwood and charcoal production may also cause local declines. Pesticide use in agricultural landscapes may reduce the insect prey base.
Conservation
This species has been recorded from the Great Limpopo Transfrontier Park and is expected to occur in many other smaller protected areas. No direct conservation measures are currently needed for this species. This species would benefit from holistic land management that reduces pesticide use and conserves buffer strips of natural vegetation to sustain insect biomass.
Recommendations for land managers and practitioners:
- Maintain buffer strips of natural vegetation and ensure natural habitat connectivity.
- Reduce pesticide use in agricultural landscapes.
Research priorities:
- Quantification of severity of local threats.
- Research is needed to determine the population size, distribution and trends (through monitoring of known subpopulations).
- Research into general ecology, habitat selection and foraging ecology.
Encouraged citizen actions:
- Citizens can assist the conservation of the species by reporting sightings on virtual museum platforms (for example, iNaturalist and MammalMAP) and therefore contribute to an understanding of the species distribution.
Bibliography
ACR. 2024. African Chiroptera Report 2024. Pretoria, South Africa.
Boyles, J.G., Cryan, P.M., McCracken, G.F. and Kunz, T.H. 2011. Economic importance of bats in agriculture. Science 332: 41–42.
Coleman, J.L., Randhawa, N., Huang, J.C.C., Kingston, T., Lee, B.P.Y., O’Keefe, J.M., Rutrough, A.L., Thong, V.D., Tsang, S.M. and Shepherd, C.R. 2024. Dying for décor: quantifying the online, ornamental trade in a distinctive bat species, Kerivoula picta. European Journal of Wildlife Research, 70(4), p.75.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An Assessment of South Africa’s Biodiversity and Ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Fahr, J. and Ebigbo, N.M. 2003. A conservation assessment of the bats of the Simandou Range, Guinea, with the first record of Myotis welwitschii (Gray, 1866) from West Africa. Acta Chiropterologica 5: 125–142.
Festa, F., Ancillotto, L., Santini, L., Pacifici, M., Rocha, R., Toshkova, N., Amorim, F., Benítez‐López, A., Domer, A., Hamidović, D. and Kramer‐Schadt, S. 2023. Bat responses to climate change: a systematic review. Biological Reviews, 98(1), pp.19-33.
GeoTerraImage. 2015. Quantifying settlement and built-up land use change in South Africa. Pretoria.
Happold, M. 2013. Myotis welwitschii. Pp 708-710. In: The Mammals of Africa. Volume IV: Hedgehogs, Shrews and Bats. Eds: M. Happold and D.C.D. Happold. Bloomsbury Publishing, London.
Hunter, S.B., Weeds, J. and Mathews, F. 2024. Monitoring the trade in bat taxidermy and specimens on e-commerce platforms. Biological Conservation, 299, p.110827.
Jacobs, D., Cotterill, F.P.D. and Taylor, P.J. 2008. Myotis welwitschii.The IUCN Red List of Threatened Species 2008.e.T14211A4422736.
Jewitt, D., Goodman, P.S., Erasmus, B.F.N., O’Connor, T.G. and Witkowski, E.T.F. 2015. Systematic land-cover change in KwaZulu-Natal, South Africa: implications for biodiversity. South African Journal of Science 111: 1–9.
Kunz, T.H., Braun de Torrez, E., Bauer, D., Lobova, T. and Fleming, T.H. 2011. Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223: 1–38.
Mammal Diversity Database. 2025. Mammal Diversity Database (Version 2.2). Zenodo. https://doi.org/10.5281/zenodo.15007505
Meester, J.A.J., Rautenbach, I.L., Dippenaar, N.J. and Baker, C.M. 1986. Classification of southern African mammals. Transvaal Museum Monographs 5: 1–359.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2020. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis. University of Witwatersrand Press, Johannesburg. 2nd edition
Monadjem, A., F. Simelane, J. T. Shapiro, B. C. Gumbi, M. L. Mamba, M. D. Sibiya, S. M. Lukhele and T. A. M. Mahlaba. 2021. Using species distribution models to gauge the completeness of the bat checklist of Eswatini. European Journal of Wildlife Research, 67: 21. doi: 10.1007/s10344-021-01463-9
Monadjem, A., Montauban, C., Webala, P.W., Laverty, T.M., Bakwo-Fils, E.M., Torrent, L., Tanshi, I., Kane, A., Rutrough, A.L., Waldien, D.L. and Taylor, P.J. 2024. African bat database: curated data of occurrences, distributions and conservation metrics for sub-Saharan bats. Scientific Data, 11(1), p.1309.
Morales, A.E., Burbrink, F.T., Segall, M., Meza, M., Munegowda, C., Webala, P.W., Patterson, B.D., Thong, V.D., Ruedi, M., Hiller, M. and Simmons, N.B. 2024. Distinct Genes with Similar Functions Underlie Convergent Evolution in Myotis Bat Ecomorphs. Molecular Biology and Evolution, 41(9), p.msae165.
Patterson, B.D., Webala, P.W., Kerbis Peterhans, J.C., Goodman, S.M., Bartonjo, M. and Demos, T.C. 2019. Genetic variation and relationships among Afrotropical species of Myotis (Chiroptera: Vespertilionidae). Journal of Mammalogy, 100(4), pp.1130-1143.
Pio, D.V., Engler, R., Linder, H.P., Monadjem, A., Cotterill, F.P., Taylor, P.J., Schoeman, M.C., Price, B.W., Villet, M.H., Eick, G. and Salamin, N. 2014. Climate change effects on animal and plant phylogenetic diversity in southern Africa. Global Change Biology, 20(5), pp.1538-1549.
Ratcliffe, J.M. 2002. Myotis welwitschi. Mammalian Species 702: 1–3.
Schoeman, C.M. and Jacobs, D.S. 2003. Support for the allotonic frequency hypothesis in an insectivorous bat community. Oecologia 134: 154–162.
Simmons, N.B. 2005. Order Chiroptera . Johns Hopkins University Press, Baltimore, USA.
Skinner, J.D. and Chimimba, C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.
Smith, R.J. et al. 2008. Designing a transfrontier conservation landscape for the Maputaland centre of endemism using biodiversity, economic and threat data. Biological Conservation 141: 2127–2138.
Taylor, P.J., Nelufule, M., Parker, D.M., Toussaint, D.C. and Weier, S.M. 2020. The Limpopo River exerts a powerful but spatially limited effect on bat communities in a semi-arid region of South Africa. Acta chiropterologica, 22(1), pp.75-86.
Weier, S.M., Linden, V.M., Hammer, A., Grass, I., Tscharntke, T. and Taylor, P.J. 2021. Bat guilds respond differently to habitat loss and fragmentation at different scales in macadamia orchards in South Africa. Agriculture, Ecosystems & Environment, 320, p.107588