

Spectacled Dormouse
Graphiurus ocularis

2025 Red list status
Near threatened
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Graphiurus ocularis – (Smith, 1829)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – GLIRIDAE – Graphiurus – ocularis
Common Names: Spectacled Dormouse, Black-and-white Dormouse, Cape Dormouse, Namtap (English), Gemsbokmuis (Afrikaans), Sepêpê (Setswana)
Synonyms: No Synonyms
Taxonomic Note: Graphiurus ocularis is thought to be closely related to G. platyops (Montgelard et al. 2003), but G. rupicola, G. monardi and G. nagtglasii were not included in the latter study. Currently, no subspecies of this dormouse have been identified, and a comprehensive overview of the molecular phylogeny of this genus is necessary (Monadjem et al. 2015).
Red List Status: NT – Near Threatened
Assessment Information
Assessors: Smith, C.1 & da Silva, J.M.2
Reviewer: Patel, T.1
Institutions: 1Endangered Wildlife Trust,2South African National Biodiversity Institute
Previous Assessors & Reviewers: Wilson, B., MacFadyen, D., Palmer, G. & Child, M.F.
Previous Contributors: Relton, C., Birss, C., Avenant, N.L., Avery, M., Baxter, R., Monadjem, A., Taylor, P. & Cassola, F.
Assessment Rationale
The Spectacled Dormouse is endemic to southwestern South Africa. While it is widely dispersed, habitat loss through agricultural expansion is suspected to have reduced area of occupancy and increased fragmentation of the population, as is evidenced by available post-2000 records occurring predominantly in protected areas. In the Western Cape alone, between 2006 and 2011, 536 km2 of land was converted to agriculture where 31% of losses occurred in Critical Biodiversity Areas. While we assume that the rocky habitats of this species are unlikely to be transformed on a large scale, the effects of climate change may make higher-elevation habitats more suitable for agriculture. Thus, the emerging threat of regional agricultural activities such as rooibos tea farming and vineyards will need to be monitored for their encroachment on Spectacled Dormouse habitat.
The species is considered rare. While the reporting frequency has been stable over the 10 years (1.2 ± 0.4 records / year) since 2005, it is 53% lower on average (2.5 ± 1.9 records / year) than the 10-year reporting frequency for the previous national assessment (1994–2004). There are also no available records at all since 2011. While it may be spurious to infer population decline based on data records because observer effort is variable, a genuine decline in the population cannot be ruled out. Thus, we list as Near Threatened A2bc given suspected population reduction over three generations due to the ongoing and emerging threats from agricultural transformation. However, we urge that further monitoring and field surveys are needed to confirm this by more accurately estimating population trends and area of occupancy. This is a precautionary listing, which should be revised as more data become available. We view this as a genuine change in status until evidence suggests otherwise.
Red List Index
Red List Index: No change
Recommended Citation: Smith C & da Silva JM. 2025. A conservation assessment of Graphiurus ocularis. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species is endemic to South Africa, where it occurs widely in the Northern Cape, Eastern Cape, and Western Cape provinces. It was first described from a locality near Plettenberg Bay, Western Cape, in 1829. There are a number of questionable records of this species (Holden 2005). For example, there is a single record from the North West Province (Skinner & Chimimba 2005), but Power (2014) cannot locate this record, and it is uncertain whether the species does occur in the province. Similarly, its occurrence in KwaZulu-Natal, due to a single skin collected in 1915, some 400 km from its known range, is doubtful (Taylor et al. 1994). Its distribution range needs to be confirmed in the northern parts of the Northern Cape and is yet to be recorded as far north as Tswalu Kalahari Reserve (D. MacFadyen unpubl. data). However, Avery and Avery (2011) reported a record from a Barn Owl (Tyto alba) pellet from Lovers Leap, 26 km north of Van Zylsrus on the Molopo River, as well as a Holocene record from the Dikbosch 1 archaeological site, 81 km west of Kimberley. Thus, the possibility that the historical range extended further north than the current range needs to be considered. It is absent from the Highveld grasslands of Gauteng and Mpumalanga (MacFadyen 2014). As in Monadjem et al. (2015), two records from north of the Gariep River (de Graaff 1981) could not be traced.
Certain areas in the Western Cape are relatively well connected and secure, and, with the advance of climate change, may well prove to be an important stronghold for the species. Within the Eastern Cape, it was only recorded at one of three sampling sites within the Sneeuberg Mountain Complex (Asante Sana Nature Reserve; Kok et al. 2012). The extent of occurrence is estimated as 344,268 km2 for all records and 42,095 km2 using just the post-2000 records. This species may qualify for a threatened listing under the B2 criterion should future research calculate area of occupancy more accurately.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
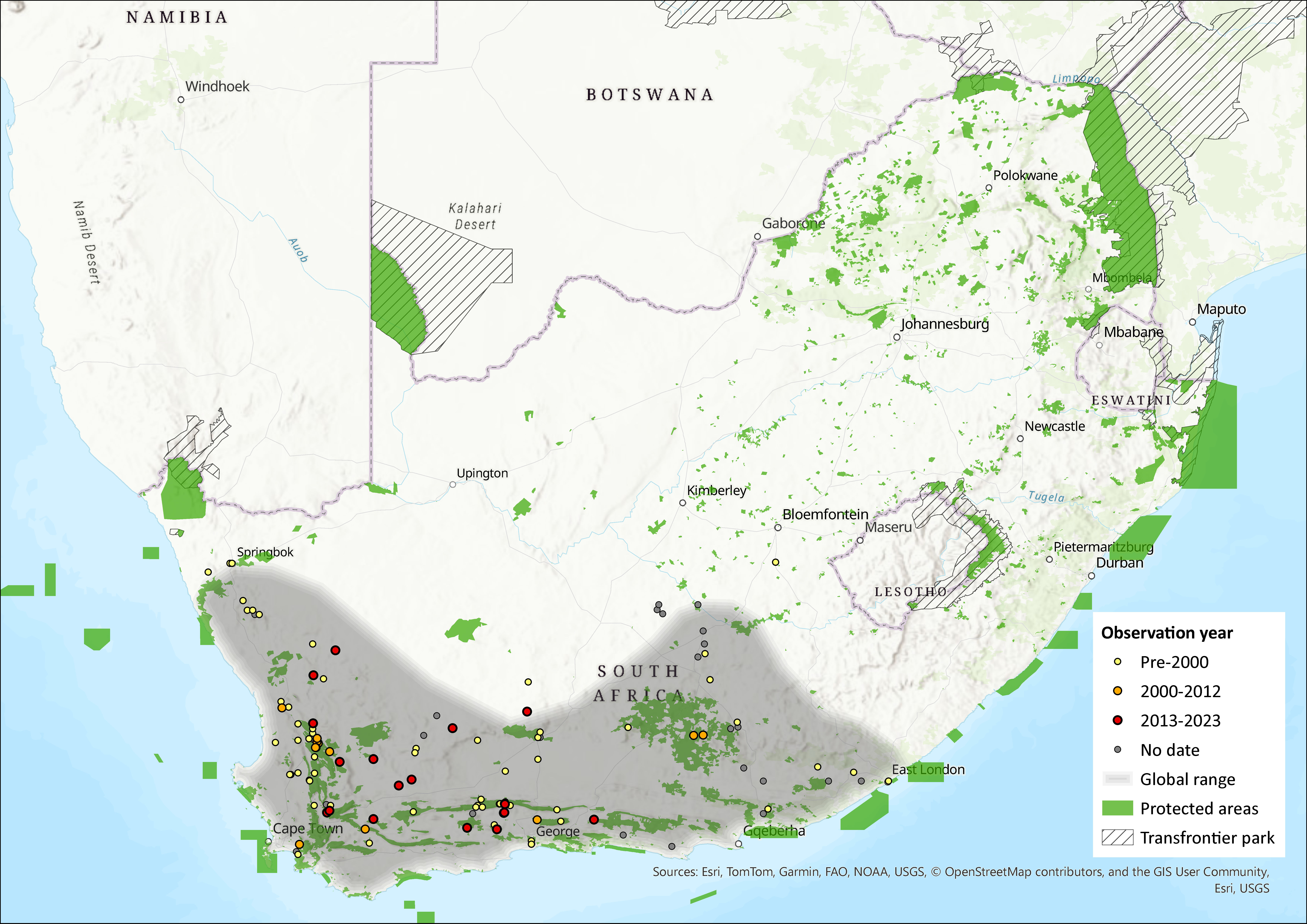
Figure 1. Distribution records for Spectacled Dormouse (Graphiurus ocularis) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| South Africa | Extant | Native | – | – |
| South Africa -> Eastern Cape Province | Extant | Native | – | – |
| South Africa -> Free State | Extant | Native | – | – |
| South Africa -> North-West Province | Extant | Native | – | – |
| South Africa -> Northern Cape Province | Extant | Native | – | – |
| South Africa -> Western Cape | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
The species occupies rocky habitats unlikely to be transformed. However, the effects of climate change may make higher-elevation habitats more suitable for agriculture which may encroach on Spectacled Dormouse habitat. For example, climate change is projected to increase the suitability of upslope habitats for viticulture, increasing the footprint of winelands by 14% by 2050 (Hannah et al. 2013). Such trends in transformation of mid- and upper slopes should be monitored. Ongoing agricultural expansion is also likely to further fragment the population.
To quantify the potential threat of climate change on the species habitat, species distribution modelling under future climate projections were performed. From a conservation perspective, the models show a consistent reduction in habitat for G. ocularis ranging between 0.56%-5% by 2040.
Population information
It is uncommon, with densities ranging between 1.8 and 3.1 animals / ha on a 7.75 ha study plot with seasonal variation in density (Channing 1984; Holden 2013). The overall population size of this species is not currently known, but it is considered uncommon to rare (Smithers 1986; Holden 2013) and population densities vary according to the suitability of the habitat. This is a relatively large animal which occurs in territories and thus is relatively easy to trap. Sampling in suitable habitats often does not reveal presence of the species. For example, trapping conducted in 2011 in Mountain Zebra National Park did not detect any individuals (Z.J.K. Madikiza & E. Do Linh San unpubl. data). Reporting rate has also declined in the Western Cape (C. Birss unpubl. data), which may be confounded by decreased observer effort. Thus, there is a priority to identify long-term monitoring sites to guide field surveys, quantify population trends and occupancy patterns, as well as to determine the effects of habitat fragmentation, which may be the cause for population discontinuity in the Cape Fold Mountains. Generation length is calculated as 2.4 years (Pacifici et al. 2013), which yields a 7.2 year three-generation period.
Population Information
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individuals in one subpopulation: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Unknown
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
While this genus has been investigated in a phylogenetic context (Montgelard et al. 2003), no population genetic assessment has been undertaken on the species.
From a genetic perspective, very little is known about this species or can be inferred. It occurs in fairly low numbers but tends to be found where suitable habitat exists. The habitat across its distribution varies, which could act to restrict populations; however, this has not been investigated. Accordingly, the species might exist as a single metapopulation or as multiple metapopulations corresponding to biomes (e.g., fynbos, Karoo, thicket). It is assumed that no extinctions have occurred within the species; hence regardless of the number of subpopulations, all are thought to exist today. Consequently, the Convention on Biological Diversity’s Global Biodiversity Framework’s complementary genetic indicator – proportion of populations maintained (PM) – would receive a value of 1.0 (all populations remaining).
While no population estimates are available, it is assumed that at least 5000 mature individuals exist across the assessment region; thereby exceeding the Ne 500 threshold (assuming an effective population size [Ne] to census size [Nc] conversion ratio between 0.1-0.3. While the presumed size is not an actual census count, the expert opinion can still be used as a proxy for quantifying the proportion of populations with an Ne > 500 indicator, which is valued at 1.0 (1/1 population with Ne > 500).
It is highly recommended that a population genetic study be undertaken to increase the available knowledge on the species and verify the estimated indicator values reported here.
Habitats and ecology
The species is associated with the sandstone formations of the Cape, which have many vertical and horizontal cracks and crevices in which to shelter and nest (Channing 1984). It has also been recorded, on camera, to visit Protea humiflora and Protea sulphurea (Kuhn et al 2017). The largest of the regional Dormouse species, this squirrel-like Dormouse with its bushy tail has a characteristic black and white face mask making it easily identifiable. It occurs within the sandstone formations of the Cape, where cracks and crevices in the rock create ideal shelters in which to nest and rest (Channing 1984), but it has also been known to use stone kraals and buildings. It is associated with rock piles, outcrops, crevices and stone kraals (Holden 2013). In Rolfontein Nature Reserve, Northern Cape Province, three specimens were caught in Setaria lindenbergiana grassland community, which is wetter than the surrounding area and covered with large boulders (Jooste & Palmer 1982). Their flattened skull allows animals to move through narrow rock crevices, and they will actively choose routes along rocks over those at ground level even though they may be considerably shorter (Holden 2013). Although this species is largely terrestrial, it is occasionally arboreal, with the type specimen (A. Smith 1829) having been captured in a tree. They are vocal and will demonstrate aggression when disturbed. In the Cederberg, they were found to occur in sympatry with Namaqua Rock Mice (Micaelamys namaquensis) and Cape Rock Sengis (Elephantulus edwardii) (Channing 1984). Similar to other species of dormouse, this species is largely an insectivore, feeding on a variety of invertebrates, but also includes birds, lizards, millipedes, spiders, scorpions and honeybees in its diet (Channing 1984).
It is nocturnal in habits, although crepuscular at times, especially on overcast days. It remains active throughout the year, but if the temperature drops sufficiently or if there is a scarcity of food, it can enter a state of torpor for up to a month (Holden 2013). In captivity it is unable to tolerate temperatures greater than 35˚C (Perrin & Ridgard 1999), and this may be a limiting factor with regards to range extensions northwards. Females are thought to produce two litters per year, 6–8 weeks apart (Channing 1984). There is some evidence that Spectacled Dormice might be territorial as pairs will occupy the same area for extended periods of time. Adults will occupy the most favourable habitats with their current offspring with less favourable habitats allocated to dispersing youngsters. Channing (1984) estimated the average lifespan to be at least 4 years.
Ecosystem and cultural services: Similar to other small mammals, this species is preyed upon by predators such as the Barn Owl (Avery et al. 2005). Additionally, this species could be used as a flagship species in the Cederberg area where it is “notorious” amongst campers for its nocturnal raiding.
IUCN Habitats Classification Scheme
|
Habitat |
Season |
Suitability |
Major Importance? |
|
3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry |
– |
Suitable |
– |
|
6. Rocky areas (eg. inland cliffs, mountain peaks) |
– |
Suitable |
– |
Life History
|
Generation Length |
Justification |
Data Quality |
|
2 |
see Pacifici et al. 2013 |
medium |
Age at maturity: female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
While there is insufficient data available to quantify severity, there is an emerging interest in the exotic pet industry for dormice despite their complicated requirements. The Spectacled Dormouse may be of particular interest due to its larger size and attractive facial features.
General notes regarding trade and use of this species: (Not specified)
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
This species is confined to rocky outcrops, and its habitat is mostly well protected. However, ongoing habitat loss (see below) and habitat fragmentation, because of plantations (such as Rooibos tea plantations) and vineyards, may impact immigration and gene flow between isolated habitats. Global climate change may further shrink its range southwards as is evidenced already by archaeological records which show they occurred significantly further north in the Holocene period (Avery & Avery 2011). Additionally, climate change is increasing the spread of alien invasive plants, which may increase the fuel load and thus the regularity and severity of wildfires (Richardson & van Wilgen 2004). This may cause habitat degradation for this species.
Current habitat trend: Possibly declining. While it occurs in rocky habitat unsuitable for agriculture, climate change may be making such higher-elevation areas more suitable for agriculture. For example, climate change is projected to increase the suitability of upslope habitats for viticulture, increasing the footprint of winelands by 14% by 2050 (Hannah et al. 2013). Such trends in transformation of mid- and upper slopes should be monitored. Ongoing agricultural expansion is also likely to further fragment the population. In the Western Cape, Pence (2014) calculated that between 2006 and 2011, 536 km2 of land was converted to agriculture (107 km2 per year), of which 31% of habitat losses occurred in Critical Biodiversity Areas. Overall, a total of approximately 2,120 km2 of natural habitat has been lost between 2000 and 2013 (G. Pence unpubl. data).
Conservation
The species occurs in several protected areas. The Cederberg Wilderness Area is particularly well known for this species, where it would appear to be at its highest density. There have also been recent records from the Jonkershoek Nature Reserve in the Boland Mountain Complex, which were, in fact, the first in 45 years. Protected area expansion to connect isolated habitats, such as the rocky sandstone habitats of the Western Cape, will presumably benefit this species. Long-term monitoring should be established to measure the effects of corridor creation.
Recommendations for land managers and practitioners:
- Ensure corridors of natural vegetation between areas of suitable habitat are protected to allow for gene flow and dispersal.
Research priorities:
- Investigate the distributional range of this species, especially, but not only, in outlier areas with suitable habitat.
- Determine population densities across this species range.
- Determine long-term population and occupancy trends inside and outside of protected areas.
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Plant indigenous trees in gardens and ensure corridors of natural vegetation remain to allow local movements.
- Clear alien invasive plant species across areas of the Cape Floristic region to reduce habitat degradation and water loss.
Bibliography
Avery DM, Avery G. 2011. Micromammals in the Northern Cape Province of South Africa, past and present. African Natural History 7:9–39.
Avery DM, Avery G, Palmer NG. 2005. Micromammalian distribution and abundance in the Western Cape Province, South Africa, as evidenced by Barn owls Tyto alba (Scopoli). Journal of Natural History 39:2047–2071.
Channing A. 1984. Ecology of the namtap Graphiurus ocularis (Rodentia: Gliridae) in the Cedarberg, South Africa. South African Journal of Zoology 19:144–149.
de Graaff G. 1981. The Rodents of Southern Africa. Butterworths, Durban, South Africa.
Hannah L, Roehrdanz PR, Ikegami M, Shepard AV, Shaw MR, Tabor G, Zhi L, Marquet PA, Hijmans RJ. 2013. Climate change, wine, and conservation. Proceedings of the National Academy of Sciences 110:6907–6912.
Holden ME. 2005. Family Gliridae. Pages 819–841 in Wilson DE, Reeder DM, editors. Mammal Species of the World. Johns Hopkins University Press, Baltimore, Maryland, USA.
Holden ME. 2013. Graphiurus ocularis Spectacled African Dormouse. Pages 128–129 in Happold DCD, editor. Mammals of Africa vol. III: Rodents, Hares and Rabbits. Bloomsbury Publishing, London, UK.
IUCN. 2016. The IUCN Red List of Threatened Species. Version 2016-3. Available at: www.iucnredlist.org. (Accessed: 07 December 2016).
IUCN. 2017. The IUCN Red List of Threatened Species. Version 2017-1. Available at: www.iucnredlist.org. (Accessed: 27 April 2017).
Jooste JF, Palmer NG. 1982. The distribution and habitat preference of some small mammals in the Rolfontein Nature Reserve. South African Journal of Wildlife Research 12:26–35.
Kok AD, Parker DM, Barker NP. 2012. Life on high: the diversity of small mammals at high altitude in South Africa. Biodiversity and Conservation 21:2823–2843.
Kühn, N., Midgley, J. and Steenhuisen, S.L., 2017. Reproductive biology of three co-occurring, primarily small-mammal pollinated Protea species (Proteaceae). South African Journal of Botany, 113, pp.337-345.
MacFadyen DN. 2014. The dynamics of small mammal populations in Rocky Highveld Grassland, Telperion, South Africa. Ph.D. Thesis. University of Pretoria, Pretoria, South Africa.
Monadjem A, Taylor PJ, Denys C, Cotterill FPD. 2015. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis. De Gruyter, Berlin, Germany.
Montgelard C, Matthee CA, Robinson TJ. 2003. Molecular systematics of dormice (Rodentia: Gliridae) and the radiation of Graphiurus in Africa. Proceedings of the Royal Society of London B: Biological Sciences 270:1947–1955.
Pacifici M, Santini L, Di Marco M, Baisero D, Francucci L, Marasini GG, Visconti P, Rondinini C. 2013. Generation length for mammals. Nature Conservation 5:89–94.
Pence GQK. 2014. Western Cape Biodiversity Framework 2014 status update: Critical Biodiversity Areas of the Western Cape. CapeNature, Cape Town, South Africa.
Perrin MR, Ridgard BW. 1999. Thermoregulation and patterns of torpor in the spectacled dormouse, Graphiurus ocularis (A. Smith 1829) (Gliridae). Tropical Zoology 12:253–266.
Power RJ. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng, South Africa.
Rathbun, G.B. (subeditor). 2005. Macroscelidea. In: J.D. Skinner and C.T. Chimimba (eds), The Mammals of the Southern African Subregion, 3rd edition, pp. 22-34. Cambridge University Press, Cambridge, UK.
Richardson DM, van Wilgen BW. 2004. Invasive alien plants in South Africa: how well do we understand the ecological impacts? South African Journal of Science 100:45–52.
Skinner JD, Chimimba CT. 2005. The Mammals of the Southern African Subregion. Third edition. Cambridge University Press, Cambridge, UK.
Smithers RHN. 1986. South African Red Data Book – Terrestrial Mammals. Page 216. Technical Report SANSP Report 125. CSIR, National Scientific Programmes Unit, South Africa.
Taylor PJ, Richardson EJ, Meester J, Wingate L. 1994. New distribution records for six small mammal species in Natal, with notes on their taxonomy and ecology. Durban Museum Novitates 19:59–66.