

Sharpe’s Grysbok
Raphicerus sharpei

2025 Red list status
Regional Population Trend
Change compared
to 2016
No Change
Overview
Raphicerus sharpei – Thomas, 1897
ANIMALIA – CHORDATA – MAMMALIA – ARTIODACTYLA – BOVIDAE – Raphicerus – sharpei
Common Names: Sharpe’s Grysbok (English), Sharpe se Grysbok, Tropiese Grysbok (Afrikaans), Isanempa (Ndebele), Phuduhudu (Setswana), Mawumbane (Swati), Pitsipitsi, Xipitsipitsi, Zipitipit (Tsonga), Grysbok de Sharpe (French), Sharpegreisbok (German)
Synonyms: No Synonyms
Taxonomic Note:
Has been considered conspecific with R. melanotis. Two subspecies proposed but validity doubtful (Hoffmann and Wilson 2013).
It was initially said that, like the Steenbok, Sharpe’s Grysbok did not have “false hooves” (a pair of supplementary hooves above the fetlocks) and was previously called the Sharpe’s Steenbok (Stevenson-Hamilton 1947; Astley Maberly 1952). It was later shown that the Sharpe’s sometimes does have false hooves but more often not (Astley Maberly 1963). Due to closer morphological links with the larger Cape Grysbok Stevenson-Hamilton 1947), although the latter always has ‘false hooves’, this species was later classified as Sharpe’s Grysbok. They thus appear to form a link between the Cape Grysbok and the Steenbok.
Red List Index: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Patel, T1 & da Silva, J.2
Reviewer: Mallon, D.3
Institutions: 1Endangered Wildlife Trust; 2South African National Biodiversity Institute; 3IUCN SSC Antelope Specialist Group
Previous Assessor: Peel, M.
Previous Reviewer: Child, M.F.
Previous Contributors: Sowry, R., Anderson, J., Joubert, D., Marshall, J., Hendry, J., Rushworth, I., Llewellyn, J., Cesare, M., Rushworth, D., Walker, A. & Kettles, R.
Assessment Rationale
This species occupies well-protected savannah habitats within the assessment region, including Kruger National Park (KNP), with dispersal possible through the Great Limpopo Transfrontier Park. The extent of occurrence is estimated to be 64,356 km2 while the area of occupancy is estimated as 27,684 km2 (for protected areas) This yields an estimated mature population size (assuming a 70% mature population structure) of 5,814-13,565 individuals, using a density estimate of 0.3–0.7 individuals / km2. Its habitat is largely protected and not expected to decline significantly in amount or quality. The conversion from livestock to wildlife ranching may instead be increasing available habitat. However, the impact of mining in the lowveld should be assessed as such activities could break habitat corridors. Available time series indicate a low-density but stable subpopulation trend over three generations (1999 to 2013) in KNP and adjacent conservancies, as well as in Save Valley Conservancy, Zimbabwe, which indicates the rate of potential immigrants will not decline. While anecdotal reports suggest that this species no longer occurs in many areas of its historical range, it may also be overlooked and under-sampled as it is difficult to detect. Although bushmeat hunting (direct or incidental) will likely cause local declines, especially outside of protected areas, there is no evidence to suggest that the population overall is in decline. Thus, we list Sharpe’s Grysbok as Least Concern. The status of Sharpe’s Grysbok is unlikely to change in the long-term if it continues to be well represented in protected areas and on private land. However, this species should be re-assessed if further monitoring data suggest a broader population decline outside protected areas.
Regional population effects: There is suspected to be dispersal and immigration across the northern border of Limpopo from Botswana and Zimbabwe through the Greater Mapungubwe Transfrontier Park and through Mozambique and Zimbabwe through the Great Limpopo Transfrontier Park. Immigration is not expected to decrease since the population in neighbouring countries is stable or increasing. However, due to its habitat preferences, described variously as thick woodland, riverine forest and broken country with bush cover (Smithers 1966), which represent threatened habitats (except for ‘broken country’), rates of immigration may be low. In the Lowveld, Sharpe’s Grysbok are mainly encountered in the ‘broken country’ type habitat (for example, along the Olifants River), which may suggest that the species is able to make use of impacted landscapes to disperse. Monitoring is needed to establish the net rescue effect and rate of dispersal/immigration.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Patel T & da Silva JM. 2025. A conservation assessment of Raphicerus sharpei. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The species occurs in savannah woodland from western and southern Tanzania (Skinner & Chimimba 2005), southwards through southeastern Democratic Republic of the Congo (Smith 1992; Skinner & Chimimba 2005), Zambia (east of the Zambezi River, absent in the north and northwest) (Smithers 1966; Smith 1992; Skinner & Chimimba 2005), Malawi (widespread) (Smithers 1966; Smith 1992; Skinner & Chimimba 2005), Mozambique (not including the coastal forested regions) (Smithers 1966; Smith 1992; Skinner & Chimimba 2005), to extreme northeastern Botswana and the eastern Caprivi Strip along the Zambezi River. The species is found in much of Zimbabwe (Smithers 1966; Smith 1992), and the Lowveld in northeast South Africa (Limpopo Province, eastern Mpumalanga) and eastern Eswatini (Stevenson-Hamilton 1947; Smithers 1966; Rautenbach 1982; Skinner & Chimimba 2005; Hoffmann & Wilson 2013). While expanding human settlements and agriculture, as well as hunting, have reduced their numbers and range, their secretive habits have enabled them to survive in pockets of their former range (Skinner & Chimimba 2005).
Within the assessment region, their stronghold is the KNP where they are most plentiful between the Olifants and Limpopo Rivers in the Mopane woodlands (Astley Maberly 1963) and the surrounding private conservancies. They also occur in the south of the park, all along the Lebombo mountain range southwards (Pienaar et al. 1987). Some specific observations follow: D. Rushworth (pers. comm. 2014) observed that some 30 years ago the strip of land on the lower slopes of the northeastern escarpment from the Swadini gate into the Blyde Nature Reserve (northwest round to the base of Manoutsa near the Strijdom Tunnel) had a viable population of Sharpe’s Grysbok. Present numbers of grysbok appear to have diminished or even disappeared on some properties (D. Rushworth pers. comm. 2014). This is supported from the area to the south of the latter observation where it appears that this species has disappeared from the farm Madrid (J. Anderson pers. comm.). Rautenbach (1982) reports a single record from the Thabazimbi district but states that it has in all likelihood disappeared from this area and the species has also disappeared from the former southeastern Transvaal (Gauteng/Mpumalanga) during recent times. Although there are no collected records, they do occur on Lapalala Wilderness in the Waterberg (J. Anderson & A. Walker pers. comm. 2014). However, Power (2014) reports that although they occur in the Waterberg, they are becoming increasingly scarce there. Stevenson-Hamilton (1947) reported that there were ‘good numbers’ of Sharpe’s Grysbok on both sides of the Lebombo Mountains between Eswatini and Mozambique and in KNP as far south as the Crocodile River at Komatipoort. This is corroborated by Skinner and Chimimba (2005), Smithers (1966), Rautenbach (1982) and Hoffmann and Wilson (2013). There are historical accounts of “grysbok” from the Drakensberg in the 1930s, 1940s and 1950s, both from Natal Parks Board rangers and forestry staff (I. Rushworth pers. comm. 2014), however, there are almost certainly no grysbok in the Drakensberg today.
The extent of occurrence (EOO) is estimated to be 64,356 km2. The EOO includes more recent records in southern Kruger, which were not present in the previous assessment. Area of occupancy was estimated to be 27,684 km2 based on protected areas using the South African Protected Areas Database of 2024 (likely to be an underestimate).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
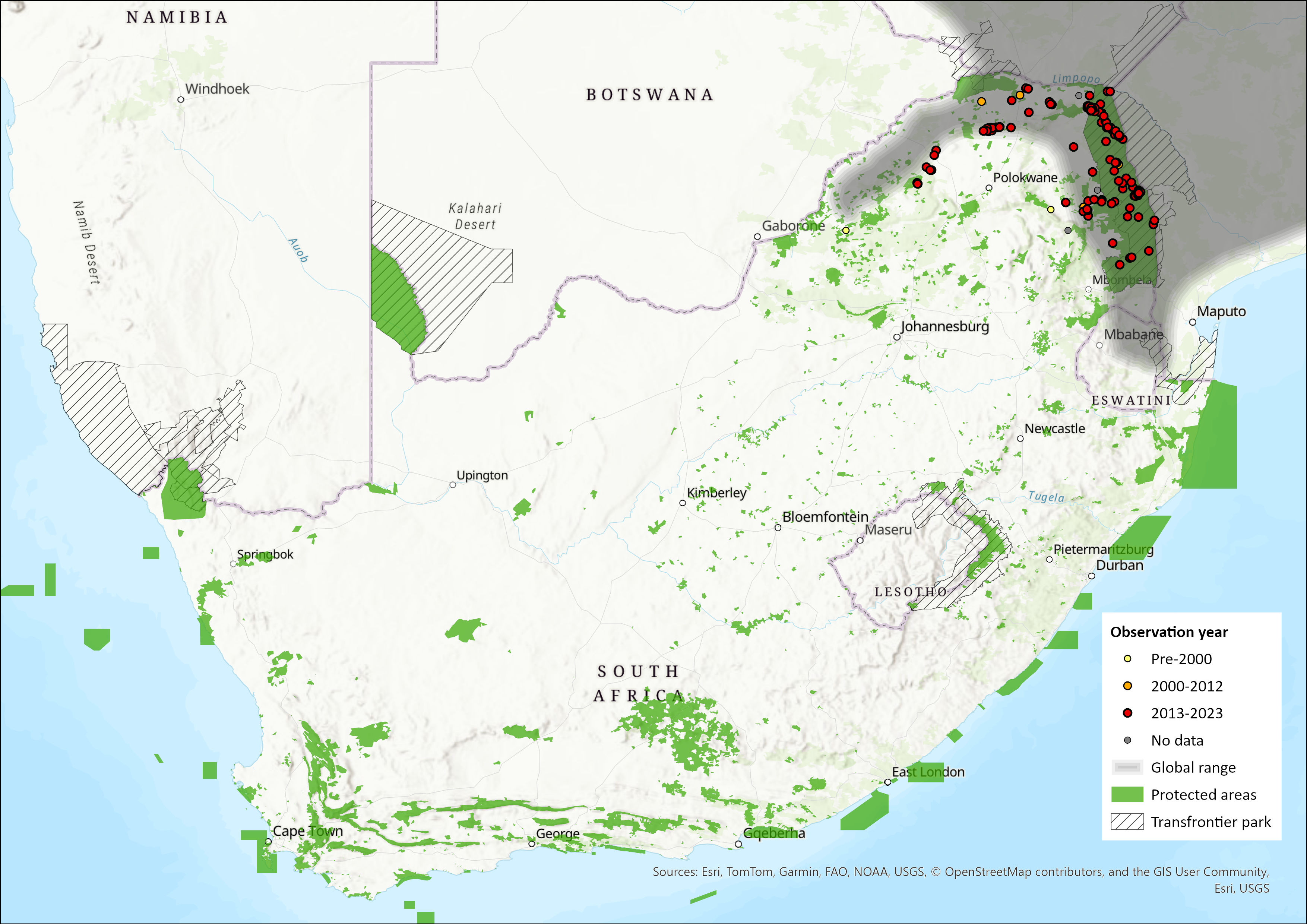
Figure 1. Distribution records for Sharpe’s Grysbok (Raphicerus sharpei) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Botswana | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia -> Caprivi Strip | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
There is no research available to indicate that Climate Change has, or will have, an impact on Sharpe’s Grysbok within the assessment region.
Population information
East (1999) summarised recorded population density estimates for this species as 0.3–0.7 animals / km², and estimated a global population size of about 95,000 animals. The 2004 national assessment estimated a population size of < 2,500 mature individuals (Friedmann & Daly 2004). Using the AOO and density estimates above, we estimate a population of 8,305-19,379 individuals, which yields a mature population size of 5,814-13,565 individuals (assuming a 70% mature population structure) for protected areas. It is thus likely that there are fewer than 10,000 mature individuals but probably not fewer than 2,500. Additionally, as Sharpe’s Grysbok are predominantly nocturnal, exceptionally shy and secretive, they can be overlooked in areas where in reality they are reasonably common so they may be more abundant than supposed (Hoffmann & Wilson 2013). For example, they are apparently common in the Soutpansberg (Power 2002).
The generation length has been calculated as 4.8 years (Pacifici et al. 2013), which yields a 14.4 year three-generation period. From the previous assessment, in protected areas and ranches where the species occurs, subpopulations were estimated to be stable. For example, data from Lapalala Wilderness indicated the regular sightings of some 5–7 animals (A. Walker unpubl. data). The Agricultural Research Institute (ARC-API) aerial survey data (1997 to 2013) indicated a low density yet stable population in the protected areas adjacent to the KNP along the Olifants River. Similarly, aerial count surveys from Selati Game Reserve revealed consistent presence of Sharpe’s Grysbok between 1 and 15 from 1998–2014. Similarly, Letaba Ranch (J. Marshall and I. Sharp unpubl. data) counted six individuals in 1995, 8 in 2003 and 14 in 2004. This number increased to 75 animals in 2012, with a steady decline over the years to 44 animals in 2018, and down to 27 in 2019 (count data from LEDET: Limpopo Economic Development, Environment and Tourism Department). Richard Sowry (pers.comm. 2014) stated that while they do not occur in any significant numbers, the subpopulation in the Kingfisherspruit section of the KNP north to the Olifants River appears to be stable.
In the Save Valley Conservancy, Zimbabwe, sightings frequency has increased between 2002 and 2010 (D. Joubert unpubl. data). Throughout its global range, the population trend is generally stable in protected areas and on private farms, but gradually decreasing elsewhere as hunting pressures increase with the growth of human populations (IUCN SSC Antelope Specialist Group 2016).
Population Information
Current population trend: Stable
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Number of mature individuals in population: 5,814-13,565
Number of mature individuals in largest subpopulation: Unknown, but most likely in KNP.
Number of Subpopulations: Unknown
Severely fragmented: No
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
The complete mitogenome for Cape and Sharpe’s Grysbok has been assembled and after comparing these with the mitogenome of R. campestris (Steenbok) it was revealed that Cape and Sharpe’s Grysboks are not sister taxa (i.e., are not most closely related to each other; de Jager and Lorenzen 2023), despite previous assumptions based on them having similar morphologies (Du Toit, 2005; Groves and Grubb 2011; Castley and Lloyd 2013).
To date, the population genetic structure for this species has not been investigated; however, historical estimates of effective population size (Ne from ~12kya) are estimated to be just under 14 000 individuals (de Jager, pers comm, June 2024). While current Ne is still under investigation using molecular tools, a proxy measure may be quantified based on the available population size estimates in the Population Section. Between 2,500-10,000 animals are thought to exist within the assessment region, therefore after applying a Ne/Nc conversion ratio of 0.1-0.3, it is estimated that Ne is between 250-3,000.
Habitats and ecology
Sharpe’s Grysbok occur in areas of scrub, thickets and grass, avoiding stands of tall grass, sometimes occupying the base of koppies and stony ridges as well as riverine vegetation (Skinner & Chimimba 2005). Specifically, they appear to be associated with Miombo (Brachystegia) woodland where there is good undercover in the form of low-growing scrub or medium-length grass (Rautenbach 1982; Skinner & Chimimba 2005; Hoffmann & Wilson 2013). Although they appear to be associated with areas of good ground cover, they are also found in pure stands of Mopane (Colophospermum mopane), and in Chobe National Park they were seen on a sandy plateau in open woodland with light grass and scrub cover (IUCN SSC Antelope Specialist Group 2016). They are predominantly browsers, but will also graze (Astley Maberly 1963; Hoffmann & Wilson 2013). They occur in areas where the majority of plant production occurs below 2 m (Jarman 1974). With territorial habits, one would expect Sharpe’s Grysbok to self-regulate their numbers. Overall, little is known about them as they are predominantly nocturnal and live in concealed habitats, often crouching low to the ground while running when flushed. Usually they occur as solitary adults, pairs or a female with a single offspring (Skinner & Chimimba 2005). Even when a pair is sharing the same bush for shade and/or shelter, they appear to lie in different parts of it (Stevenson-Hamilton 1947).
Locally, in the protected areas adjacent to the KNP (eastern Lowveld), they are found predominantly in the Lowveld Rugged Mopaneveld (Astley Maberly 1963; Mucina & Rutherford 2006). This area has dissected steep slopes with dense shrub cover interspersed with large trees and a moderate herbaceous layer (Skinner & Chimimba 2005; Mucina & Rutherford 2006). There is a subpopulation in the Letaba Ranch reserve (J. Marshall pers. comm. 2014), which is in agreement with Astley-Maberley (1963), and, further east, they are seen in the Makuleke between the Luvuvhu and Limpopo Rivers. In the Kingfisherspruit section of the KNP, the species is seen mostly north of the Timbavati River in the Mopane/Combretum veld (R. Sowry pers. comm. 2014). This concurs with J. Llewllyn (pers. comm. 2014) who reports most of the sightings in the Umbabat from the farms Sibon and Buffelsbed, which are to the north of Richard Sowry’s section. West of the latter reserves, M. Cesare (pers. comm. 2014) reports that they are relatively common in the hilly Commiphora woodland adjacent to the Olifants in the Balule Nature Reserve. Still further west it appears that there are small but stable subpopulations on the Selati (D. Joubert pers. comm. 2014) and Makalali Game Reserves (R. Kettles pers. comm. 2014). In the eastern Lowveld along the Olifants River and west towards the Drakensberg Mountains, this species occupies a niche in the shallow-soiled, broken, stony terrain just below the rocky outcrop zone (Stevenson-Hamilton 1947; Astley Maberly 1963; Potgieter et al. 1971; Werger 1978; Rautenbach 1982; Skinner & Chimimba 2005; M. Cesare pers. comm. 2014; I. Rushworth pers. comm. 2014). The terrain is undulating, broken and stony with mainly shallow, sandy soils and appears to be prime habitat for grysbok. A detailed description of these habitats in the protected areas adjacent to the KNP is presented in Peel et al. (2007) and in the KNP in Gertenbach (1983). Sharpe’s Grysbok are also found in a diversity of other habitats from dry sandy veld in Botswana, Miombo woodland in Zambia and Zimbabwe and in riverine areas of Hwange (Smithers 1966, 1971).
Ecosystem and cultural services: None recorded
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 3.4. Shrubland -> Shrubland – Temperate | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species is poached as bushmeat, which may cause local subpopulation declines, especially outside protected areas. They are also hunted as trophies (Fiorenza 1983), and thus grysbok are commercially valuable to hunting and live sale, where they are a sought after trophy and have become a highly valued species (D. Rushworth pers. comm. 2014).
In addition to protection in provincial reserves and national parks, private protected areas such as those adjacent to the west of the KNP have generally had a positive effect on this species. This is due to relatively lighter stocking densities in the areas in which this species occurs. Overgrazing would probably lead to declines in the numbers of this species.
| Subsistence: | Rationale: | Local Commercial: | Further detail including information on economic value if available: |
| Yes | Bushmeat hunting | Yes | Trophy hunting and live animal sales |
National Commercial Value: No
International Commercial Value: No
| End Use | Subsistence | National | International | Other (please specify) |
| 1. Food – human | true | – | – | – |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: No known ranching of this species. No known captive breeding of this species.
Threats
There are no major threats, although they have been eliminated from some parts of their former range by the spread of settlement and agriculture, and associated habitat destruction, and hunting for meat (IUCN SSC Antelope Specialist Group 2016). Within the assessment region, human settlements and mining along the edge of KNP have similarly reduced habitat and habitat quality and may have broken habitat corridors. This may facilitate bushmeat hunting, which may cause local declines, especially as human density increases along protected area edges (Wittemyer et al. 2008). Its secretive habits, however, may enable it to withstand considerable hunting pressure. Despite this, subpopulations may be gradually decreasing outside of protected areas as bushmeat poaching increases with the growth of human populations and this should be monitored.
The Sharpe’s Grysbok is water independent and excessive artificial water will reduce groundcover around waterholes and increase Impala (Aepyceros melampus) numbers, which may sustain a denser Leopard (Panthera pardus) population. This could affect long-term density of prey such as Grysbok (M. Cesare pers. comm. 2014; J. Llewellyn pers. comm. 2014). Similarly, the increase in mesocarnivores, such as Black-backed Jackal (Lupulella mesomelas) and Caracal (Caracal caracal), may have a detrimental impact on the population in the long-term (D. Rushworth, R. Kettles pers. comm. 2014). In the vicinity of the Olifants River, increasing Baboon (Papio ursinus) troops that inhabit the same zone poses the greatest threat to the species young and in turn this could lead to declines in the future (M. Cesare pers. comm. 2014).
Increased stocking rates of livestock and/or larger game species reduce grass and ground cover required for Sharpe’s Grysbok survival. A knock-on effect of this is the reduction of seasonal wild fires that resulted in a mature woody layer with less low coppice for lateral cover and browse (D. Rushworth pers. comm. 2014). Similarly, there is an observed general increase in the density of the shorter woody layer in the lowveld areas which, with a decline in the frequency of fires, should increase the amount of suitable habitat for this species.
Current habitat trend: Stable. Savannah habits are well protected in the assessment region (Driver et al. 2012). For example, habitat is generally well conserved in protected areas adjacent to KNP as there is little human settlement. Further, the increase in the size of the wildlife ranching estate should ensure the security of the population in the longer term. However, in some areas, such as along the Olifants River, mining will certainly have had a negative impact on the habitat that Sharpe’s occupy. The extent of this degraded area is still fortunately small. It is thought that overgrazing by livestock and/or cattle would have a detrimental effect on the survival of this species. Increases in large ungulate numbers have resulted in a reduction of grass and ground cover, which is a critical habitat for this species (Skinner & Chimimba 2005). On the other hand there is an observed increase in the density of the woody layer in the lowveld areas which, with a decline in the frequency of fires, should increase the amount of suitable habitat for this species.
Conservation
About one third of the total population occurs in protected areas (IUCN SSC Antelope Specialist Group 2016), the largest one within the assessment region being KNP and Associated Private Nature Reserves (APNR). However, they also occur on a number of smaller formally and privately protected areas within the region, including wildlife ranches. There are currently no interventions necessary for this species. However, private landowners are encouraged to create conservancies to conserve suitable habitat for this species. Additionally, the regulation of translocation is required to prevent mixing of ecotypes and hybridisation with Cape Grysbok. There is a substantial distance between the southernmost occurrence of Sharpe’s Grysbok and the northeasternmost occurrence of the Cape Grysbok (Hoffmann & Wilson 2013). Further field surveys and long-term monitoring is needed to establish more accurate range maps and population trends.
Recommendations for land managers and practitioners:
- Maintenance of the integrity of the habitat of the Sharpe’s Grysbok. This includes ensuring that fragmentation of habitat does not occur and that protected areas remain intact (for example: Letaba Ranch along the Letaba River eastward; the area to the west of the R40 moving eastwards into the APNR and KNP into the Lebombo mountains along the Olifants River; and the area between the Luvuvhu and Limpopo Rivers).
- Regulate translocation to avoid hybridisation with Cape Grysbok.
- Long term monitoring of the species is needed to assess population trend.
Research priorities: Very little is known about the Sharpe’s Grysbok other than that they are largely nocturnal, generally solitary, and occurring at low densities. Studies on their ecology would be useful and, due to the relatively wide variety of habitats in which they occur (see above), studies would need to be quite broad within the sub-region. Specific research priorities include:
- Density estimates and population size calculation across its range.
- Information on occurrence outside protected areas.
- Effects of wildlife ranching on the species.
- Assessing the need for metapopulation management in the medium term to ensure genetic diversity (J. Anderson pers. comm. 2014).
- Similarly, studies documenting reintroduction efficacy of ‘surplus’ animals into previously occupied or suitable areas
- Monitoring is needed to establish the net rescue effect and rate of dispersal/immigration.
Encouraged citizen actions:
- Landowners should drop internal fences to form conservancies.
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
Bibliography
Astley Maberly CT. 1952. Animals of the Kruger National Park. A.C. White, Bloemfontein, South Africa.
Astley Maberly CT. 1963. The Game Animals of Southern Africa. Nelson. Cape and Transvaal Printers, Cape Town, South Africa.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., Van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An assessment of South Africa’s biodiversity and ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
East, R. (compiler). 1999. African Antelope Database 1998. IUCN, Gland, Switzerland and Cambridge, UK.
Fiorenza P. 1983. Encyclopaedia of Big Game Animals in Africa with Their Trophies. Larousse and Company, New York, USA.
Friedmann, Y. and Daly, B. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. Conservation Breeding Specialist Group (SSC/IUCN) and Endangered Wildlife Trust, Parkview, South Africa.
Gertenbach WPD. 1983. Landscapes of the Kruger National Park. Koedoe 26: 9-121.
Hoffmann, M. and Wilson, V. J. 2013. Raphicerus sharpei Sharpe’s Grysbok. In: J. S. Kingdon and M. Hoffmann (eds), The Mammals of Africa, pp. 308-310. Bloomsbury, London, UK.
IUCN SSC Antelope Specialist Group. 2016. Raphicerus sharpei. The IUCN Red List of Threatened Species 2016: e.T19307A50193414
Jarman PJ. 1974. The social organisation of antelope in relation to their ecology. Behaviour 48: 215-267.
Mucina, L. and Rutherford, M.C. 2006. The Vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria, South Africa.
Pacifici, M., Santini, L., Di Marco, M., Baisero, D., Francucci, L., Grottolo Marasini, G., Visconti, P. and Rondinini, C. 2013. Generation length for mammals. Nature Conservation 5: 87–94.
Peel MJS, Kruger JM, MacFadyen S. 2007. Woody vegetation of a mosaic of protected areas adjacent to the Kruger National Park South Africa. Journal of Vegetation Science 18: 807-814.
Pienaar, U. de V., Joubert, S. C. J., Hall-Martin, A., de Graff, G. and Rautenbach, I. L. 1987. Field guide to the mammals of the Kruger National Park. Struik and the National Parks Board of Trustees, Cape Town and Pretoria.
Potgieter DJ, Skaife SH, du Plessis PC. 1971. Animal Life in Southern Africa. NASOU Limited, South Africa.
Power RJ. 2002. Prey selection of leopards Panthera pardus in the Soutpansberg, Limpopo Province, and the utilisation options for this population. Mammal Research Institute, University of Pretoria, Pretoria.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Rautenbach, I. L. 1982. Mammals of the Transvaal. Ecoplan monograph, N.S. Supplemento 1: 111-211.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Smith SJ. 1992. Large Mammals of Africa. Rowland Ward Publications, Johannesburg and San Antonio, USA.
Smithers RHN. 1966. The Mammals of Rhodesia, Zambia and Malawi. Collins, London, UK.
Smithers, R.H.N. 1971. The mammals of Botswana. National Museums of Rhodesia, Museum Memoir 4: 1-340.
Stevenson-Hamilton J. 1947. Wild Life in South Africa. Cassell, London, UK.
Werger MJA. 1978. Biogeography and Ecology of Southern Africa. Junk Publishers, The Hague, Netherlands.
Wittemyer, G., Elsen, P., Bean, W.T., Burton, A.C.O. and Brashares, J.S. 2008. Accelerated human population growth at protected area edges. Science 321: 123-126.