

Schlieffen’s Bat
Nycticeinops schlieffenii

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Nycticeinops schlieffenii – (Peters, 1859)
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – VESPERTILIONIDAE – Nycticeinops – schlieffeni
Common Names: Schlieffen’s Bat, Schlieffen’s Serotoe, Schlieffen’s Twilight Bat (English), Schlieffen se Vlermuis, Schlieffen-vlermuis Klein Dakvlermuis (Afrikaans)
Synonyms: Nycticeius schlieffeni (Peters, 1859); N[ycticejus] adovanus (von Heuglin, 1877) [not used as valid]; Scotophilus minimus (Noack, 1887); Scotophilus schlieffeni (Yerbury & O. Thomas, 1895) [incorrect subsequent spelling]; Scoteinus schlieffeni albiventer (O. Thomas & Wroughton, 1908); Scoteinus schlieffeni australis (O. Thomas & Wroughton, 1908); Scoteinus schlieffeni bedouin (O. Thomas & Wroughton, 1908); Nycticeius africanus (G. M. Allen, 1911); Scotoecus cinnamomeus (von Wettstein, 1916); Scoteinus schlieffeni fitzsimonsi (A. Roberts, 1932)
Taxonomic Note:
The genus has been characterised by taxonomic uncertainty; with the phylogenetic placement of some species a source of debate amongst authors (Hill and Harrison 1987; Monadjem et al. 2013, Decher et al. 2015; Hutterer et al. 2019; Monadjem et al. 2021; Demos et al. 2024). The same is true of Nycticeinops schlieffeni. This species was originally listed under the genus Nycticeius and later classified to Nycticeinops (Harrison and Hill 1987). The validity of the subspecies Nycticeinops schlieffeni australis (Thomas & Wroughton 1908) and N. s. fitzsimonsi (Roberts 1932) is currently considered uncertain (Monadjem et al. 2020; Happold 2013). The northern Namibian population appears isolated and hence might represent a unique evolutionary lineage, but further molecular studies are needed to resolve this (Monadjem et al. 2010). Torrent et al. (2025) described two distinct mitochondrial genetic lineages – N. schlieffeni from West Africa and the Arabian Peninsula and N. cf. schlieffeni from East Africa. The study did not include samples from the assessment region; hence the taxonomic affinity of such individuals remains uncertain.
Red List Status: LC – Least Concern (IUCN version 3.1)
Assessment Information
Assessors: Richards L.R.1, Naidoo, T.1, Balona, J.2 & da Silva J.M.3
Reviewer: Smith, C.4
Institutions: 1Durban Natural Science Museum, 2Gauteng and Northern Regions Bat Interest Group, 3South African National Biodiversity Institute, 4Endangered Wildlife Trust
Previous Assessors and Reviewers: Monadjem, A., Jacobs, D., Cohen, L., MacEwan, K., Richards, L.R., Schoeman, C., Sethusa, T. & Taylor, P.
Previous Contributors: Relton, C., Child, M.F. & Raimondo, D.
Assessment Rationale
The species is listed as of Least Concern in view of its wide distribution in Africa as well as in the assessment region where the extent of occurrence (EOO) is 210,087 km2. This species is common, occurs in protected areas and has no known major threats.
Regional population effects: The resident N. schlieffeni population appears to be continuous with those of Mozambique and Zimbabwe through the Great Limpopo Transfrontier Park and the Greater Mapungubwe Transfrontier Conservation Area. However, it has low wing-loading (Norberg & Rayner 1987) so rescue effects are uncertain.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Richards LR, Naidoo T, Balona J & da Silva JM. 2025. A conservation assessment of Nycticeinops schlieffenii. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species has a widespread but disjunct distribution (Happold 2013). Its distribution is split into two major groups: one range in West Africa, including north Senegal, south Mauritania, Burkina Faso, Togo, Benin, Nigeria and south Angola (ACR 2024), with an additional isolated population in northern Namibia (Monadjem et al. 2020); and one covering much of eastern and southern Africa (as far south as northeastern Eswatini, in the Mlawula Nature Reserve, and northern KwaZulu-Natal). Recent molecular studies provide evidence for genetic distinction between East and West African populations (Torrent et al. 2025, see population genetics). The species is absent from much of the Horn of Africa, northeastern Ethiopia, eastern Tanzania, and northeastern Mozambique. In southern Africa, it occurs widely in the eastern and northern parts of the region but is absent from the arid west and from the tropical forests of the extreme north (Monadjem et al. 2020). Within the assessment region, the species is restricted to the northeastern regions, recorded from Limpopo, Mpumalanga and KwaZulu-Natal provinces of South Africa, as well as in Eswatini. The estimated extent of occurrence is 210,087 km². Its range is continuous with Zimbabwe, Zambia, Mozambique, Malawi and southern Democratic Republic of the Congo (Monadjem et al. 2020).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 3 m asl (Monadjem et al. 2024)
Elevation Upper Limit (in metres above sea level): 2,922 m asl (Monadjem et al. 2024)
Depth Lower Limit (in metres below sea level): N/A
Depth Upper Limit (in metres below sea level): N/A
Depth Zone: N/A
Map
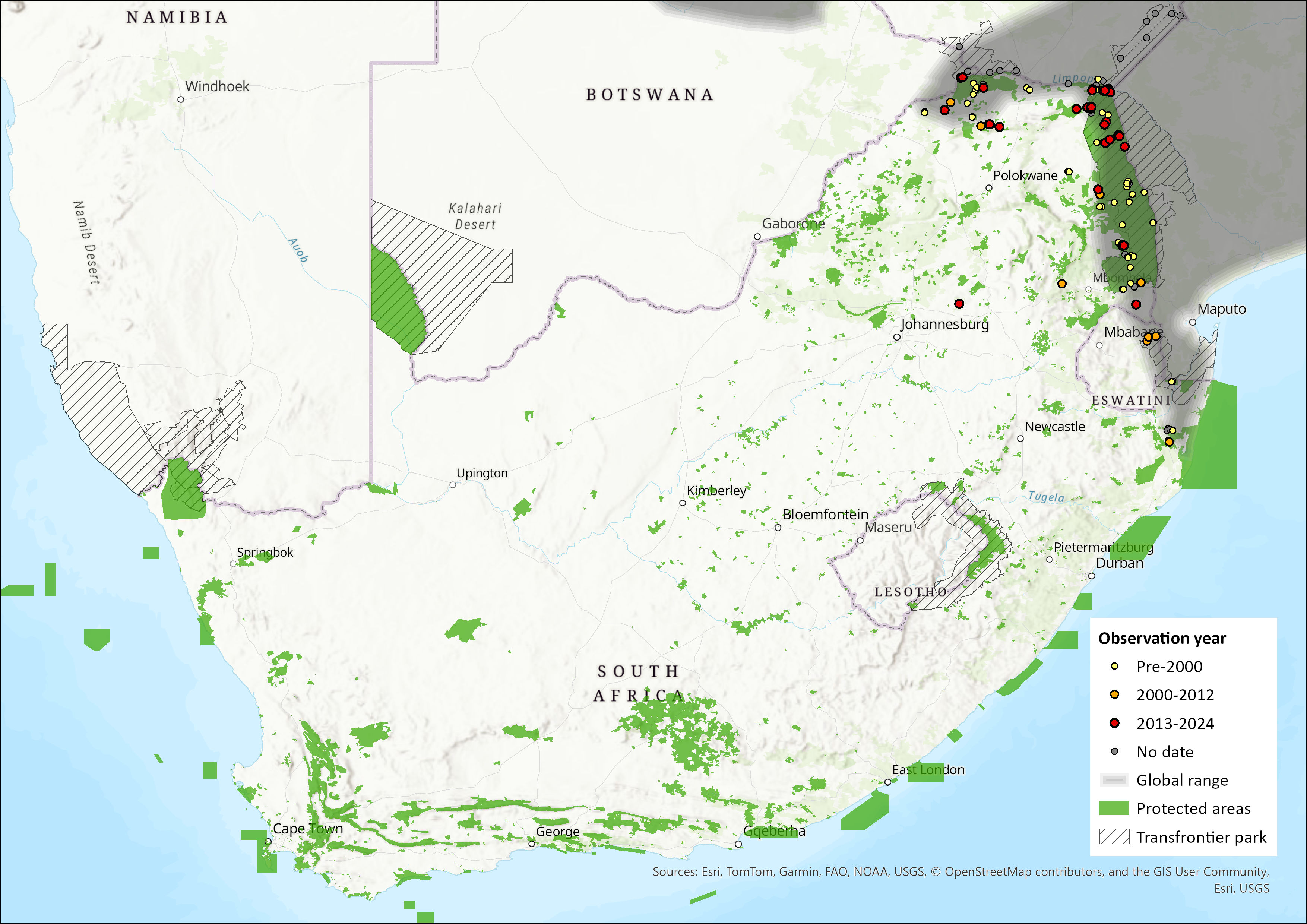 Figure 1. Distribution records for Schlieffen’s Bat (Nycticeinops schlieffenii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Schlieffen’s Bat (Nycticeinops schlieffenii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical, Palearctic
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Benin | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Burkina Faso | Extant | Native | – | – |
| Cameroon | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Chad | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Djibouti | Extant | Native | – | – |
| Egypt | Extant | Native | – | – |
| Eritrea | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Ethiopia | Extant | Native | – | – |
| Ghana | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mali | Extant | Native | – | – |
| Mauritania | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Niger | Extant | Native | – | – |
| Nigeria | Extant | Native | – | – |
| Saudi Arabia | Extant | Native | – | – |
| Senegal | Extant | Native | – | – |
| Somalia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Togo | Extant | Native | – | – |
| Uganda | Extant | Native | ||
| Yemen | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: N/A
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
No formal studies have investigated the potential impact of climate change on the species within the assessment region. Studies on captive bats predicted a very high urine concentrating ability (4562 mOsmol/kg) (Happold and Happold 1988), suggestive of an increased dehydration tolerance (Geluso 1978). However, the effects of climate change and physiological responses of bats are severely lacking (Festa et al. 2023), and focussed investigations are necessary to determine species-specific responses to the phenomenon.
Population Information
This gregarious species is considered common within its range in northeastern South Africa and Eswatini; and individuals are known to roost together in large numbers (Rosevear 1965; Rautenbach 1982). Similarly, they are often very numerous in mopane woodland in northern Zimbabwe (F. P. D. Cotterill unpubl. data). They are well represented in museums, with over 300 specimens examined in Monadjem et al. (2010).
Current population trend: Stable
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individuals in one subpopulation: Uncertain; see population genetics section
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Uncertain; see population genetics section
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
Demos et al. (2024), based on nuclear intron analyses proposed that all currently recognised species within Nycticeinops, with the exception of N. schlieffeni, should be reclassified under the genus Afropipistrellus. Recent analyses by Torrent et al. (2025) identified two distinct genetic lineages N. schlieffeni (N. schlieffeni from Senegal and Yemen) and another tentatively assigned as N. cf. schlieffeni (Kenya) are sister to the genus Afropipistrellus. The phylogenetic affinities of southern African populations remain undetermined due to the lack of representative samples from the subregion in the analyses.
No population genetic studies have been conducted on this species; however, it is expected that animals within the assessment region form part of a larger metapopulation with neighbouring countries. And given their common status, it is expected that the effective population size for this metapopulation, exceeds the threshold of 500 individuals to ensure the maintenance of genetic diversity.
Habitats and ecology
The species occurs across a range of habitats, including dry and moist savannah (miombo and mopane woodlands), semi-desert grassland and shrubland, riparian forest (but avoids true forests), Acacia (Vachellia)–Commiphora bushland, some coastal woodlands and forests, but avoids most arid areas with the exception of areas with persistent riverine habitats (Happold 2013). It appears to be closely associated with low-lying savannahs, where it may be abundant in well-wooded places such as riparian vegetation along rivers and drainage lines (Monadjem & Reside 2008). The species has a diverse roosting ecology, utilising both natural and man-made roosts. It roosts by day in hollow branches, crevices in hollow trees, rock crevices, roofs and cellars (Skinner & Chimimba 2005; Monadjem et al. 2020; Happold 2013). They were also recorded from under the bark of a dead Senegalia (previously Acacia) nigrescens tree in mopane savannah in Zinave National Park (Smithers & Lobão-Tello 1976). In the assessment region, the species is recorded from the Mopane, Lowveld and Central Bushveld bioregions.
Considered a clutter-edge forager, it is characterised by broad wings with corresponding low wing-loading to allow for slow, yet manoeuvrable flight (Norberg and Rayner 19877; Monadjem et al. 2020). This insectivorous species feeds predominantly on aerial insects such as Coleoptera, Diptera, Lepidoptera, Trichoptera and Hymenoptera (Fenton et al. 1977a, 1998; Fenton & Thomas 1980; Aldridge & Rautenbach 1987). Vegetative matter may also constitute a proportion of their diet (Seamark & Bogdanowicz 2002).
Ecosystem and cultural services: As this species is insectivorous, it may contribute to controlling insect populations that damage crops (Boyles et al. 2011; Kunz et al. 2011). Ensuring a healthy population of insectivorous bats can thus decrease the need for pesticides.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 8.1. Desert -> Desert – Hot | – | Suitable | – |
Life History
Generation Length: Unknown
Age at Maturity: Female or unspecified: Unknown
Age at Maturity: Male: Unknown
Size at Maturity (in cms): Female: Mean forearm length = 3.11 ± 0.12 cm; mean total length = 7.94 ± 0.58 cm
Size at Maturity (in cms): Male: Mean forearm length = 3.13 ± 0.11 cm; mean total length = 7.58 ± 0.34 cm
Longevity: Unknown
Average Reproductive Age: Unknown
Maximum Size (in cms): Females: maximum forearm length = 3.30 cm; Males = 3.36 cm
Size at Birth (in cms): Unknown
Gestation Time: Approximately 11 weeks (van der Merwe and Rautenbach 1986)
Reproductive Periodicity: Seasonal monoestry, with births coinciding with the austral summer months (November to December) (van der Merwe and Rautenbach 1986)
Average Annual Fecundity or Litter Size: 2-3 young born per litter (van der Merwe and Rautenbach 1986)
Natural Mortality: predated upon by bat hawks (Fenton et al. 1977b)
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Unknown
Congregatory: The species is known to roost singly, in pairs, as well as small and large colonies in the subregion (Rautenbach 1982; Smithers and Lobão Tello 1976). Individuals are known to congregate during foraging bouts (Smithers 1983).
Systems
System: Terrestrial
General Use and Trade Information
There is no evidence to suggest that this species is traded or harvested within the assessment region.
Local Livelihood: None reported
National Commercial Value: Likely to contribute to ecosystem services, in the form of pest insect population control where it occurs in agroecosystems.
International Commercial Value: Likely to assist with insect population control in respect of exported produce.
End Use: None reported
Is there harvest from captive/cultivated sources of this species? No
Harvest Trend Comments: N/A
Threats
No major threats have been identified for this wide-ranging species within the assessment region. However, roost disturbance and removal through logging of large trees for fuelwood and charcoal production may lead to local declines. For example, recent land-cover analysis reveals that 20% of forest and woodland cover was lost from 1990 to 2006 in the Soutpansberg Mountain region due to logging, residential expansion and pine/eucalyptus plantations (Munyati & Kabanda 2009). In general, climate change has been identified as an increasing global threat to other bat species as it amplifies water scarcity issues and negatively affects primary plant production and prey availability (Sherwin et al. 2013) and may similarly impact N. schlieffeni.
Conservation
This species occurs within a number of protected areas within the assessment region, including Great Limpopo Transfrontier Park, Mlawula Nature Reserve (Eswatini), Hans Merensky Nature Reserve, Baobab Tree Reserve, Ndumo Game Reserve and iSimangaliso Wetland Park. No specific conservation measures are deemed necessary at present. However, the protection of large trees required for roosting and conservation of riparian habitats, particularly in more arid regions, will benefit this species. Public education concerning the essential ecosystem services that insectivorous bats render is key to their conservation.
Recommendations for land managers and practitioners:
- Reduce pesticide use in agricultural landscapes.
- Protect and preserve large trees, which provide valuable roosting sites for this species.
- Protection of riparian habitats in water scarce regions.
Research priorities:
- Inclusion of samples from the assessment region in further molecular DNA and taxonomic revision studies, particularly with regards to the recent description of genetically distinct lineages within this widespread species (Torrent et al. 2025).
- Targeted surveys, inclusive of passive acoustic monitoring, in under sampled areas of the subregion to document additional occurrence sites.
- Increased sampling, inclusive of passive acoustic monitoring, throughout the predicted species range in the assessment region (see Monadjem et al. 2010).
- Investigations into the potential impacts of climate change, and associated increasing temperatures, varying rainfall patterns, and extreme weather events, on the biogeography and physiology of the species.
- Studies investigating the severity of potential threats on the local population.
Encouraged citizen actions:
- Citizens can assist the conservation of the species by reporting sightings on virtual museum platforms (for example, iNaturalist and MammalMAP) and therefore contribute to an understanding of the species distribution.
- Report any grounded bats to your nearest bat interest group and lodge specimens with a natural science collection institute.
Bibliography
ACR. 2024. African Chiroptera Report. 2024. AfricanBats NPC, Pretoria. i – xv; 1 – 1397.
Aldridge HDJN, Rautenbach IL. 1987. Morphology, echolocation and resource partitioning in insectivorous bats. The Journal of Animal Ecology 56: 763–778.
Boyles, J.G., Cryan, P.M., McCracken, G.F. and Kunz, T.H. 2011. Economic importance of bats in agriculture. Science 332: 41–42.
Demos, T.C., Webala, P.W., Monadjem, A. and Patterson, B.D. 2024. Nuclear introns support the subtribe Laephotina and recently proposed genera of African Vespertilionidae. Acta Chiropterologica 26:143-152
Driver, A., Sink, K.J., Nel, J.N., Holness, S., van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An Assessment of South Africa’s Biodiversity and Ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Fenton, M.B. et al. 1998. Bats and the loss of tree canopy in African woodlands. Conservation Biology 12: 399–407.
Fenton, M.B., Boyle, N.G.H., Harrison, T.M. and Oxley, D.J. 1977a. Activity patterns, habitat use, and prey selection by some African insectivorous bats. Biotropica 9: 73–85.
Fenton, M.B., Cumming, D.H.M. and Oxley, D.J. 1977b. Prey of bat hawks and availability of bats. The Condor, 79(4), pp.495-497.
Fenton, M.B. and Thomas, D.W. 1980. Dry-season overlap in activity patterns, habitat use, and prey selection by sympatric African insectivorous bats. Biotropica 12: 81–90.
Festa, F., Ancillotto, L., Santini, L., Pacifici, M., Rocha, R., Toshkova, N., Amorim, F., Benítez‐López, A., Domer, A., Hamidović, D. and Kramer‐Schadt, S. 2023. Bat responses to climate change: a systematic review. Biological Reviews, 98(1), pp.19-33.
Geluso, K.N. 1978. Urine concentrating ability and renal structure of insectivorous bats. Journal of Mammalogy, 59(2), pp.312-323.
Happold, M. 2013. Nycticeinops schlieffeni Schlieffen’s Twilight Bat. Pages 595–597 in Happold M, Happold DCD, editors. Mammals of Africa. Volume IV: Hedgehogs, Shrews and Bats. Bloomsbury Publishing, London, UK.
Happold, D.C.D. and Happold, M.J.J.Z. 1988. Renal form and function in relation to the ecology of bats (Chiroptera) from Malawi, Central Africa. Journal of Zoology, 215(4), pp.629-655.
Kunz, T.H., Braun de Torrez, E., Bauer, D., Lobova, T. and Fleming, T.H. 2011. Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223: 1–38.
Monadjem, A. and Reside, A. 2008. The influence of riparian vegetation on the distribution and abundance of bats in an African savanna. Acta Chiropterologica 10: 339–348.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2010. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis. University of Witwatersrand Press, Johannesburg.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2020. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis, Second Edition. University of Witwatersrand Press, Johannesburg.
Monadjem, A., Montauban, C., Webala, P.W., Laverty, T.M., Bakwo-Fils, E.M., Torrent, L., Tanshi, I., Kane, A., Rutrough, A.L., Waldien, D.L. and Taylor, P.J. 2024. African bat database: curated data of occurrences, distributions and conservation metrics for sub-Saharan bats. Scientific Data, 11(1), p.1309.
Munyati, C. and Kabanda, T.A. 2009. Using multitemporal Landsat TM imagery to establish land use pressure induced trends in forest and woodland cover in sections of the Soutpansberg Mountains of Venda region, Limpopo Province, South Africa. Regional Environmental Change 9: 41–56.
Norberg, U.M. and Rayner, J.M. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society B: Biological Sciences 316: 335–427.
Rautenbach, I.L. 1982. Mammals of the Transvaal. Mammals of the Transvaal. No. 1, Ecoplan Monograph.
Rosevear, D.R. 1965. The Bats of West Africa. British Museum (Natural History), London, UK.
Seamark, E.C.J. and Bogdanowicz, W. 2002. Feeding ecology of the common slit-faced bat (Nycteris thebaica) in KwaZulu-Natal, South Africa. Acta Chiropterologica 4: 49–54.
Sherwin, H.A., Montgomery, W.I. and Lundy, M.G. 2013. The impact and implications of climate change for bats. Mammal Review, 43(3), pp.171-182.
Skinner, J.D. and Chimimba, C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.
Smithers, R.H.N. 1983. The Mammals of the Southern African Subregion. University of Pretoria Press, Pretoria. 734 pp.
Smithers, R.H.N. and Lobão-Tello, J.L.P. 1976. Check list and atlas of the mammals of Mozambique. Trustees of the National Museums and Monuments of Rhodesia, Salisbury, Rhodesia.
Torrent, L., Juste, J., Garin, I., Aihartza, J., Dalton, D.L., Mamba, M., Tanshi, I., Powell, L.L., Padidar, S., Mudarra, J.L.G., Richards, L. and Monadjem, A. 2025. Taxonomic revision of African pipistrelle-like bats with a new species from the West Congolean rainforest. Zoological Journal of the Linnean Society 204 (2): DOI:10.1093/zoolinnean/zlaf020
Van der Merwe, M. and Rautenbach, I.L. 1986. Multiple births in Schlieffen’s bat, Nycticeius schlieffenii (Peters, 1859)(Chiroptera: Vespertilionidae) from the southern African subregion. African Zoology, 21(1), pp.48-50.