

Rusty Bat
Pipistrellus rusticus

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Pipistrellusrusticus – (Tomes, 1861)
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – VESPERTILIONIDAE – Pipistrellus – rusticus
Common Names: Rusty Pipistrelle, Rusty Pipistrelle Bat, Rusty Bat, Rustic Pipistrelle (English), Roeskleurvlermuis (Afrikaans)
Synonyms: Scotophilus rusticus 1861; Pipistrellus marrensis Thomas and Hinton 1923; Eptesicops rusticus Roberts 1926
Taxonomic Note:
Recent molecular DNA studies suggest that the southern African individuals are more closely related to Pipistrellus cf. hesperidus from Senegal than Kenyan P. hesperidus (Torrent et al. 2025). The species can sometimes be distinguished from other similarly sized pipistrelles in the field by its bicoloured fur, which features rusty-tinged tips and pale rusty-grey bases (Monadjem et al., 2020). Males are readily identifiable from other pipistrelle-like bats due to their distinctive penile morphology (Fasel et al. 2020). Two subspecies were recognised (Simmons 2005): P. r. marrensis occurring in Senegal to Ethiopia and Kenya; and the slightly larger P. r. rusticus in southern Africa (Zambia, Namibia, Botswana, Zimbabwe and South Africa). The validity of these subspecies is uncertain.
Red List Status: LC – Least Concern (IUCN version 3.1)
Assessment Information
Assessors: Richards, L.R.1 & da Silva, J.M.2
Reviewer: Balona, J.3
Institutions: 1Durban Natural Science Museum, 2South African National Biodiversity Institute, 3Gauteng and Northern Regions Bat Interest Group
Previous Assessors and Reviewers: Monadjem, A., Jacobs, D., Cohen, L., MacEwan, K., Richards, L.R., Schoeman, C., Sethusa, T. & Taylor, P.
Previous Contributors: Relton, C., Raimondo, D. & Nicholson, S.K.
Assessment Rationale
The species is listed as Least Concern in view of its wide distribution within the assessment region (estimated extent of occurrence is 245,363 km2), its occurrence in multiple protected areas (including Great Limpopo Transfrontier Park and Greater Mapungubwe Transfrontier Conservation Area) and because there are no major identified threats that could cause widespread population decline. It occurs primarily in savannah woodland habitats that are well protected within the assessment region (Monadjem et al. 2020).
Regional population effects: It is likely that the range of resident populations is continuous with those of Zimbabwe and Mozambique through its occurrence in transfrontier conservation areas. However, it has low wing loading (Norberg & Rayner 1987) and thus recruitment and rescue effects are uncertain.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Richards LR & da Silva JM. 2025. A conservation assessment of Pipistrellus rusticus. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species has a broad, yet patchy distribution through sub-Saharan Africa. Westerly records extend to Senegal and the Gambia, while, in East Africa, specimens have been collected from Ethiopia near the border of Somalia. Records from central Africa are limited to southern Chad and the northern region of the Central African Republic (ACR 2024). The southern portion of their range extends as far as the northern and northeastern reaches of South Africa and central Namibia, and they are known to occur in Zimbabwe and Mozambique, with an isolated population in Malawi (Monadjem et al. 2020).
Within the assessment region, it occurs in the northern provinces of South Africa; namely: Limpopo, Mpumalanga, Gauteng and the North West, as well as Eswatini (Monadjem et al. 2020). Ecological niche models suggest a broader potential distribution for the species across the eastern regions of the assessment area, predominantly within grassland and savannah biomes, while largely excluding low-lying coastal habitats (Monadjem et al. 2010). Estimated extent of occurrence in the assessment region is 245,363 km2.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 6 m asl (Monadjem et al 2024)
Elevation Upper Limit (in metres above sea level): 2,527 m asl (Monadjem et al 2024)
Depth Lower Limit (in metres below sea level): N/A
Depth Upper Limit (in metres below sea level): N/A
Depth Zone: N/A
Map
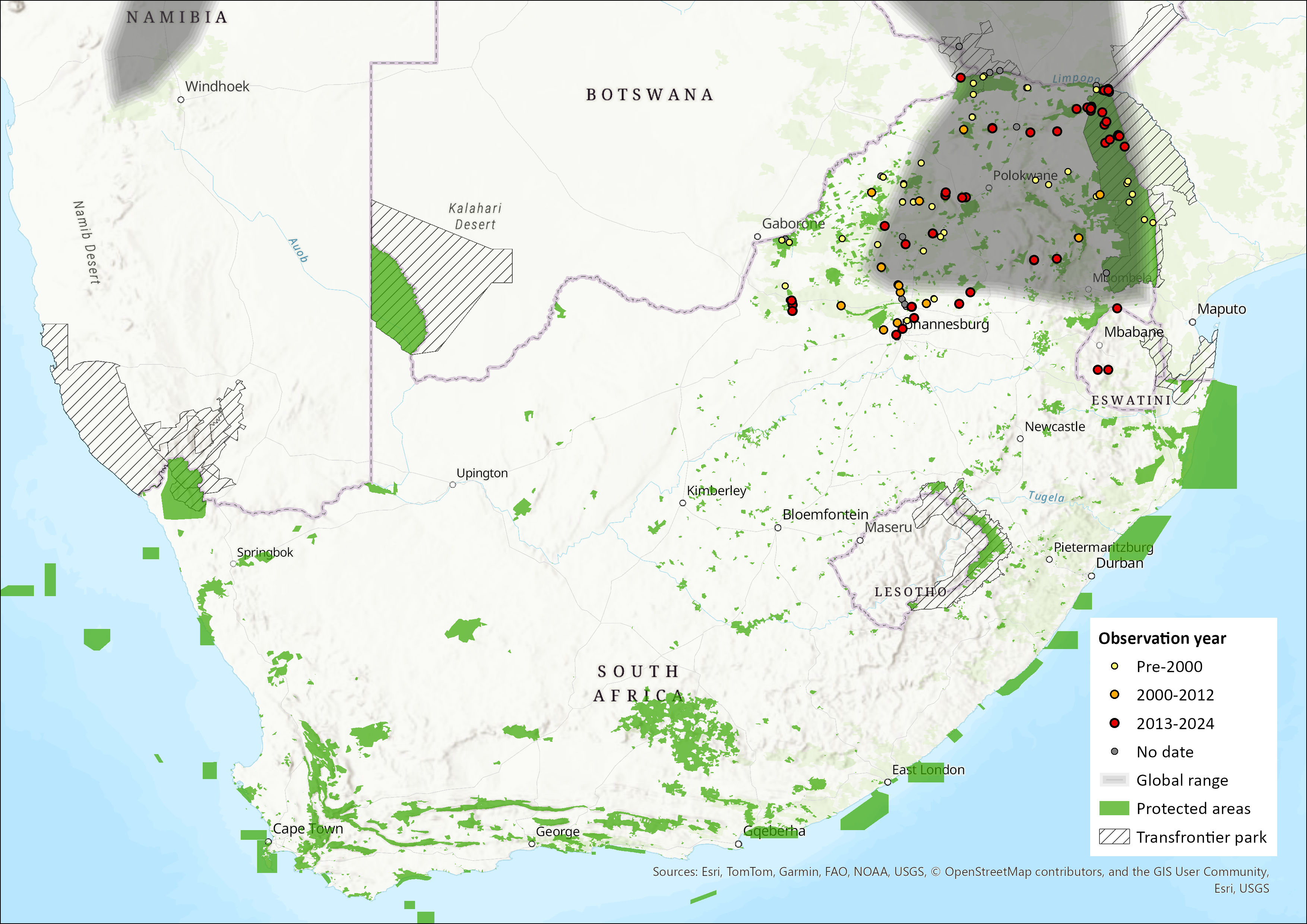 Figure 1. Distribution records for Rusty Pipistrelle (Pipistrellus rusticus) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Rusty Pipistrelle (Pipistrellus rusticus) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Burkina Faso | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Chad | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Ethiopia | Extant | Native | – | – |
| Gambia | Extant | Native | – | – |
| Ghana | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Nigeria | Extant | Native | – | – |
| Senegal | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: N/A
FAO Area Occurrence
FAO Marine Areas: (N/A
Climate change
No empirical studies have been directed towards the potential impacts of climate change on the species. However, predictive modelling has identified precipitation seasonality as the primary factor influencing its distribution (Babiker & Salata 2012). Rising temperatures, combined with increasingly erratic rainfall patterns and heightened seasonality, may significantly alter the species’ distribution within the assessment region.
Population Information
Although this species has a patchy distribution, it is considered locally common (Van der Merwe and Rautenbach 1990; ACR 2024). For example, in the Limpopo valley, it is locally common in mopane woodland where rocky habitat (Clarens sandstones) is also present (Monadjem et al. 2020). It is well represented in museums, with over 150 specimens examined in Monadjem et al. (2020). Further monitoring is required to estimate population size and trends. Currently, we assume the species is stable based on it being sampled in recent field surveys (Taylor et al. 2013; Power 2014; Brinkley et al. 2021; de Jong et al. 2025; Gumede et al. 2025).
Current population trend: Stable
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individuals in one subpopulation: Assumed to be one population within the assessment region, however population genetic studies are needed to verify this.
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: None
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: While not certain, it seems improbable.
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: While not certain, it seems improbable.
Probability of extinction in the wild within 100 years: While not certain, it seems improbable.
Population Genetics
No population genetic study has been conducted on this species; however, based on its known distribution it is expected to exist as a single metapopulation within the assessment region and adjacent countries. Despite being considered common, population size estimates are unknown; however, given that it is likely part of a large metapopulation it is likely that the effective population size exceeds 500. A population genetic investigation would be needed to confirm this genetic structure and diversity.
Habitats and ecology
This species inhabits savanna woodland ecosystems, where it exhibits a strong association with water sources (Monadjem et al. 2020). It is, however, absent from both mesic miombo woodlands and xeric savanna environments (Monadjem et al. 2020). It has been captured along the riparian fringes of the Wilge River in a heterogenous grassland-dominated landscape (de Jong et al. 2025). There are limited data available for the roosting habits of this species, but they are commonly netted over water and have been collected in tree crevices and hollows (Roberts 1951; Taylor 2000), under the bark of dead Vachellia (previously Acacia) trees (Rautenbach 1982), and roof spaces in old buildings (Skinner & Chimimba 2005). It is a clutter-edge forager with a diet consisting of Coleoptera, Diptera and Lepidoptera (Aldridge and Rautenbach 1987). There is slight sexual dimorphism; with the females being slightly heavier than their male counterparts (van der Merwe & Rautenbach 1990; Monadjem et al. 2020). Studies into the reproductive ecology of the Rusty Bat in Limpopo revealed that copulation occurs in April, followed by a period of sperm storage in the female until August and September, when ovulation and fertilization take place (van der Merwe and Rautenbach 1990).
Ecosystem and cultural services: This species has been recorded from the margins of macadamia agroecosystems (Weier et al. 2021). As this species is insectivorous, it may contribute to controlling insect populations that damage crops (Boyles et al. 2011; Kunz et al. 2011). Ensuring a healthy population of insectivorous bats can thus decrease the need for pesticides.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Suitable | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Unknown | – |
Life History
Generation Length: Unknown
Age at Maturity: Female or unspecified: Unknown
Age at Maturity: Male: Unknown
Size at Maturity (in cms): Female: Mean forearm length = 2.91 ± 0.11 cm (Monadjem et al. 2020)
Size at Maturity (in cms): Male: Mean forearm length = 2.87 ± 0.11 cm (Monadjem et al. 2020)
Longevity: Unknown
Average Reproductive Age: (Not specified)
Maximum Size (in cms): Female maximum forearm length = 3.12 cm; Male maximum forearm length = 2.99 cm (Monadjem et al. 2020)
Size at Birth (in cms): (Not specified)
Gestation Time: approximately 3-4 months (van der Merwe and Mostert 2005)
Reproductive Periodicity: Exhibit seasonal monoestry, with copulation in April, fertilisation in August and birthing occurring in November (van der Merwe and Mostert 2005).
Average Annual Fecundity or Litter Size: Females give birth to two pups each season (van der Merwe and Mostert 2005)
Natural Mortality: Malan et al. (2022) documented predation by bat hawks (Macheiramphus alcinus).
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Unknown
Congregatory: Individuals within the assessment region may roost in small colonies; such behavior has been documented in tree-roosting colonies in Zambia (Taylor 2000).
Systems
System: Terrestrial
General Use and Trade Information
There is no evidence to suggest that this species is traded or harvested within the assessment region.
|
Subsistence: |
Rationale: |
Local Commercial: |
Further detail including information on economic value if available: |
|
Yes |
– |
– |
– |
National Commercial Value: Likely to contribute to overall insect pest population suppression in agroecosystems such as macadamia orchards.
International Commercial Value: Likely to contribute to overall insect pest population suppression in agroecosystems associated with exported produce such as macadamia.
End Use: None reported.
Is there harvest from captive/cultivated sources of this species? N/A
Harvest Trend Comments: N/A
Threats
No major threats have been identified for this species. Ongoing habitat loss from agricultural transformation and the use of pesticides in agricultural landscapes may reduce the insect prey base for this species. Given the species’ strong association with water bodies and riparian habitats, water pollution is likely to pose a significant threat – both to the species itself and to the insect communities it relies on for food.
Conservation
Within the assessment region the species is protected in more than 10 conservation areas, including large reserves Great Limpopo Transfrontier Park, Greater Mapungubwe Transfrontier Conservation Area and Madikwe Nature Reserve. No direct conservation measures are currently needed. However, outside of protected areas, it would benefit from holistic land management that reduces pesticide use, conserves water bodies and riparian habitats, and conserves buffer strips of natural vegetation to sustain insect biomass.
Recommendations for land managers and practitioners:
- Reduce pesticide use in agricultural landscapes and maintain buffer strips of natural vegetation.
- Conservation of water bodies and associated habitats.
Research priorities:
- Molecular genetic studies, inclusive of population-level investigations, are required to determine the taxonomic status of previously described subspecies.
- Studies into its roosting behaviour, habitat selection and population status are needed.
- Investigations into the possible threats faced by this species.
Encouraged citizen actions:
- Citizens can assist the conservation of the species by reporting sightings on virtual museum platforms (for example, iNaturalist and MammalMAP) and therefore contribute to an understanding of the species distribution. However, this species can be difficult to distinguish by untrained professionals in the field from other pipistrelle species (Kearney 2005), so professional verification will be necessary.
Bibliography
ACR. 2024. African Chiroptera Report. 2024. AfricanBats NPC, Pretoria. i – xv; 1 – 1397.
Aldridge, H.D.J.N. and Rautenbach, I.L. 1987. Morphology, echolocation and resource partitioning in insectivorous bats. The Journal of Animal Ecology, pp.763-778.
Babiker Salata, H. A. 2012. Environmental factors influencing the distribution of bats (Chiroptera) in South Africa. PhD Thesis, University of Cape Town: i – xix; 1 – 303. URL: https://open.uct.ac.za/handle/11427/6887.
Boyles, J.G., Cryan, P.M., McCracken, G.F. and Kunz, T.H. 2011. Economic importance of bats in agriculture. Science 332: 41–42.
De Jong, G.J., Melville, H.I. and Richards, L.R. 2024. The insectivorous bat assemblage in Telperion Nature Reserve, Mpumalanga, South Africa: species inventory and call library. Acta Chiropterologica, 26(2), pp.315-327. Driver, A., Sink, K.J., Nel, J.N., Holness, S., van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An Assessment of South Africa’s Biodiversity and Ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Fasel, N.J., Mamba, M.L. and Monadjem, A. 2020. Penis morphology facilitates identification of cryptic African bat species. Journal of Mammalogy, 101(5), pp.1392-1399.
Gumede, L., Comley, J., Schmitt, M.H., Stears, K. and Parker, D.M. 2025. Using Acoustic Monitoring to Assess Insectivorous Bat Richness and Activity in a Sub‐Tropical Savanna. Biotropica, 57(5), p.e70082.
Kearney, T.C., Volleth, M., Contrafatto, G. and Taylor, P.J. 2002. Systematic implications of chromosome GTG-band and bacula morphology for southern African Eptesicus and Pipistrellus and several other species of Vespertilioninae (Chiroptera: Vespertilionidae). Acta Chiropterologica 4: 55–76.
Kearney, T.C. 2005. Systematic revision of southern African species in the genera Eptesicus, Neoromicia, Hypsugo and Pipistrellus (Chiroptera: Vespertilionidae). Ph.D. Thesis, University of KwaZulu-Natal.
Kunz, T.H., Braun de Torrez, E., Bauer, D., Lobova, T. and Fleming, T.H. 2011. Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223: 1–38.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2010. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis. University of Witwatersrand Press, Johannesburg.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2020. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis, Second Edition. University of Witwatersrand Press, Johannesburg.
Monadjem, A., Montauban, C., Webala, P.W., Laverty, T.M., Bakwo-Fils, E.M., Torrent, L., Tanshi, I., Kane, A., Rutrough, A.L., Waldien, D.L. and Taylor, P.J. 2024. African bat database: curated data of occurrences, distributions and conservation metrics for sub-Saharan bats. Scientific Data, 11(1), p.1309.
Norberg, U.M. and Rayner, J.M. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society B: Biological Sciences 316: 335–427.
Power, R.J. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng, South Africa.
Rautenbach, I.L. 1982. Mammals of the Transvaal. Ecoplan Monograph, No. 1, Pretoria.
Roberts, A. 1951. The Mammals of South Africa. The Trustees of the Mammals of South Africa. Central News Agency, Johannesburg, South Africa.
Simmons, N.B. 2005. Order Chiroptera . Johns Hopkins University Press, Baltimore, USA.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Taylor, P.J., Sowler, S., Schoeman, M.C. and Monadjem, A. 2013. Diversity of bats in the Soutpansberg and Blouberg Mountains of northern South Africa: complementarity of acoustic and non-acoustic survey methods. South African Journal of Wildlife Research 43: 12–26.
Taylor, P.J. 2000. Bats of Southern Africa: Guide to Biology, Identification, and Conservation. University of Natal Press, Pietermaritzburg, South Africa.
Taylor, P.J., Strydom, E., Richards, L., Markotter, W., Toussaint, D.C., Kearney, T., Cotterill, F.P.D., Howard, A., Weier, S.M., Keith, M. and Neef, G. 2022. Integrative taxonomic analysis of new collections from the central Angolan highlands resolves the taxonomy of African pipistrelloid bats on a continental scale. Zoological Journal of the Linnean Society, 196(4), pp.1570-1590.
Torrent, L., Juste, J., Garin, I., Aihartza, J., Dalton, D.L., Mamba, M., Tanshi, I., Powell, L.L., Padidar, S., Garcia Mudarra, J.L. and Richards, L., Monadjem, A. 2025. Taxonomic revision of African pipistrelle-like bats with a new species from the West Congolean rainforest. Zoological Journal of the Linnean Society, 204(2), p.zlaf020. van der Merwe, M. and Rautenbach, I.L. 1990. Reproduction in the rusty bat, Pipistrellus rusticus, in the northern Transvaal bushveld, South Africa. Journal of Reproduction and Fertility 89: 537–542.
van der Merwe, M. & Mostert, T. 2005. Ovarian activity and early embryonic development in the rusty bat, Pipistrellus rusticus. African Zoology, 40(1), 45–54.
Weier, S.M., Linden, V.M., Hammer, A., Grass, I., Tscharntke, T. and Taylor, P.J. 2021. Bat guilds respond differently to habitat loss and fragmentation at different scales in macadamia orchards in South Africa. Agriculture, Ecosystems & Environment, 320, p.107588.