

Red Veld Rat
Aethomys chrysophilus

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Aethomys chrysophilus – (de Winton, 1897)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – MURIDAE – Aethomys -chrysophilus
Common Names: Red Veld Rat, Red Rock Rat, Red Veld Aethomys (English)
Synonyms: acticola, alticola, imago, singidae, voi (Linzey et al. 2013)
Taxonomic Note: African veld rats (genus Aethomys) are a diverse group of murid rodents that are geographically associated with East, Central and southern Africa. Molecular data (e.g. Ducroz et al. 2001; Lecompte et al. 2008; Aghová et al. 2018) supported the inclusion of Aethomys in the tribe Arvichanthini. More recent phylogenomic analyses (Mikula et al. 2021) placed Aethomys as a sister genus to the monophyletic group of arboreal rodents (Thallomys, Grammomys and Thamnomys). Nine species have been recognised within Aethomys (Monadjem et al. 2015; Wilson et al. 2017) but morphological and genomic data have shown that A. strannarius is a synonym of A. alghazal (previously known as A. hindei) and A. ineptus is just an intraspecific lineage of A. chrysophilus (Brosseau-Acquaviva et al. unpubl.) although different in chromosome number (A. ineptus is 2n = 44 while A. chrysophilus is 2n = 50; Chimimba et al. 1999; Castiglia et al. 2003). Aethomys ineptus is therefore considered here a younger synonym of A. chrysophilus (Josef Bryja pers. Comm.).
Red List Status: LC – Least Concern
Assessment Information
Assessors: Russo, I.M.1 & da Silva, J.M.2
Reviewer: Bryja, J.3
Contributor: Patel, T.4
Institutions:1Cardiff University, 2South African National Biodiversity Institute,3Institute of Vertebrate Biology CAS, Brno, Czechia,4Endangered Wildlife Trust
Previous Assessors & Reviewers: Linzey, A. V., MacFadyen, D., Russo, I. M., Taylor, P. & Bryja, J.
Previous Contributors: Child, M.F., Avenant, N., Avery, M., Baxter, R., Monadjem, A., Palmer, G. & Wilson, B.
Assessment Rationale
Red Veld Rats have a wide distribution in the Zambezian biogeographical region of Africa (sensu Linder et al. 2012), occurring at the southern limits of their range within the assessment region. They occur in Kruger National Park (north of the Olifants River) and Kgalagadi Transfrontier Park and can also occur in agricultural or disturbed habitats. No widespread decline is expected and thus the Least Concern listing is retained. However, as this species relies on ground cover, overgrazing and imprudent fire management could cause local declines and should be avoided. Further research is needed to more accurately delineate the relative distributions of this species..
Regional population effects: Dispersal is possible along connected habitats of the northern border of South Africa (Limpopo, North West and Northern Cape Provinces), Namibia, Botswana, Zimbabwe and Mozambique.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Russo IM & da Silva JM. 2025. A conservation assessment of Aethomys chrysophilus. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species is widely distributed in the savanna habitats of southern and East Africa, being found from southern Kenya to northern South Africa (along the Limpopo Valley in the Limpopo Province). Specifically, it occurs in Namibia, excluding the western desert regions, and is widely distributed throughout Zimbabwe (Skinner and Chimimba 2005). The species has also been positively identified in Francistown, Botswana (Russo et al. 2006) and the Chimanimani National Park (Mamba et al. 2024) in Mozambique which corresponds with the distributional range proposed by Linzey et al. (2003) in northern Botswana and southern/central Mozambique. The most important phylogeographic structure is the Kafue-Zambezi River system that divides all mitochondrial sequences into two main groups (Mazoch et al. 2018).
Within the assessment region specifically, A. chrysophilus occurs in a relatively narrow band bordering Botswana, Zimbabwe, and Mozambique in the Limpopo, North West and (possibly) Northern Cape Provinces (Linzey and Chimimba 2008, Linzey et al. 2013). In the west, the range extends southward to 24°S (near Rooibokkraal in the North West Province) and in the east to 24°S in the vicinity of the Olifants River in Kruger National Park (Linzey and Chimimba 2008), while its southern distributional range limit is roughly correlated with the southern distribution of mopane (Colophospermum mopane) and baobab (Adansonia digitata) trees (Linzey and Chimimba 2008). It is also expected to occur in the northern bushveld in the lower Marico River valley of the North West Province (Power 2014) and was recorded in low numbers on the northern plains of the Kruger National Park (MacFadyen 2007).
Aethomys chrysophilus is primarily found at elevations < 1,000 m asl (Chimimba and Linzey 2008).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
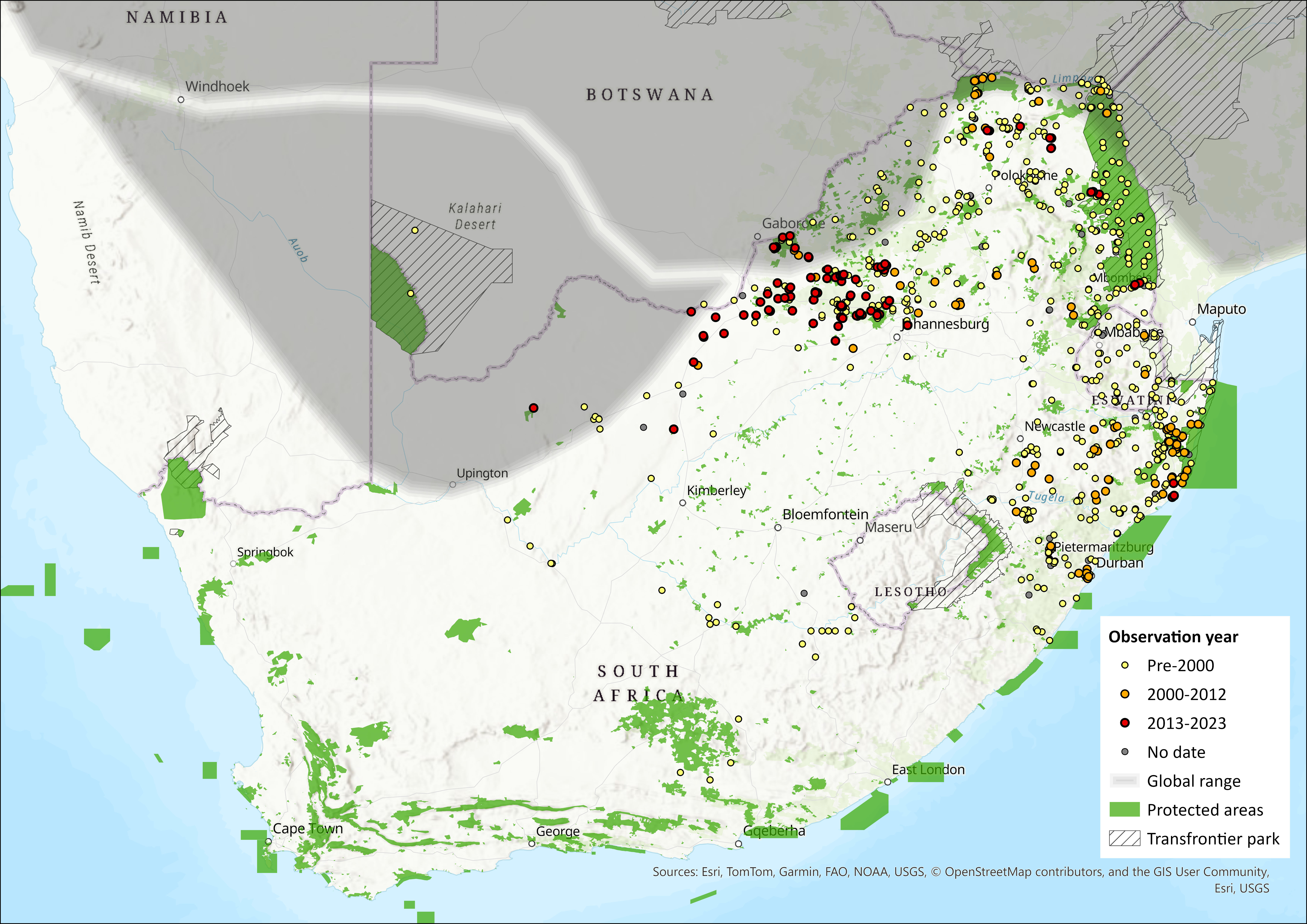
Figure 1. Distribution records for Red Veld Rat (Aethomys chrysophilus) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Lesotho | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
No recent research or literature has focused on how climate change might affect this species in the future.
Population
Studies indicate that the Red Veld Rat is generally a low-density (< 8 individuals/ha) species (Linzey et al. 2013) that attains highest numbers in either the hot-wet or early cool-dry seasons, and is least abundant in the late hot-dry season, with densities up to 6.2 individuals/ha (Linzey and Kesner 1997a, Linzey and Chimimba 2008). The species typically comprises a secondary component of small mammal communities, often being the least abundant among commonly occurring species, comprising between 2% and 15% of the community abundance (Linzey and Kesner 1997b, Linzey and Chimimba 2008). Densities are also highest in areas with good ground cover of either vegetation or rocks.
Population Information
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Current population trend: Stable
Continuing decline in mature individuals: No
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: No, savannah habitats largely intact and the species can occur in altered agricultural landscapes.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
A phylogeographic study by Mazoch et al. (2018) identified seven parapatric clades for A. chrysophilus based on mitochondrial cytb and nuclear sequencing data; two clades of which were present within the assessment region, one classified as A. ineptus. However, the placement of A. ineptus within the A. chrysophilus phylogeny and the lack of support in cranial morphology, suggests A. ineptus should be collapsed into A. chrysophilus (J. Bryja, unpublished data). Despite the imminent collapse of A. ineptus, sufficient genetic evidence exists to recognize two genetically distinct subpopulations within the assessment region, which are primarily separated by the Limpopo River (but are in contact now and exchange their genes).
Based on the available information, at least two genetic subpopulations (possibly ESUs given differences in karyotype) of A. chrysophilus occur within the assessment region. From this, it is possible to quantify one of the two population genetic indicators in the Convention on Biological Diversity’s Global Biodiversity Framework’s (GBF). The complementary genetic indicator – proportion of populations maintained (PM), would receive a value of 1.0 (2/2 populations maintained).
Habitats and ecology
A.chrysophilusis a habitat generalist occupying a variety of savanna woodlands (Linzey et al. 2013), with the specific requirement being good ground cover in the form of rocky crevices, piles of boulders and debris, fallen trees or holes in termitaria, thorn fences around agricultural lands, thick scrub or clumps of grass and forbs (Skinner and Chimimba 2005, Linzey and Chimimba 2008, Linzey et al. 2013). These rats also may be recorded in the vicinity of homesteads and barns. They are found throughout savanna–woodland habitats that include varying combinations of grass–herbaceous ground cover, shrub understory, and miombo (Brachystegia ) or mopane woodlands (Davis 1962). This species dominated the more pristine habitats characterised by rock cover at Maguga Dam, Eswatini (Avenant and Kuyler 2002). However, it has also been recorded from Aristida grasslands (adjacent to dwellings, crop lands and roads), specifically Ngongoni Three-awn Grass (Aristida junciformis), an indicator of past disturbance, which demonstrates its ability to exist in disturbed areas (Fuller and Perrin 2001). It is absent from arid regions or high-elevation forested habitats. Frequently found in association with Micaelamys namaquensis but is more abundant when the latter is absent (Linzey et al. 2013).
The Red Veld Rat is nocturnal and digs burrows in soil in areas with a good grass cover, as well as constructing nests in rock niches, bases of trees and termite mounds (Linzey and Chimimba 2008). It is terrestrial but climbs trees occasionally (Linzey et al. 2013). It is omnivorous but typically relies more on plant than animal foods.
Ecosystem and cultural services: They act as seed dispersers and are a good forage species for carnivores.
IUCN Habitats Classification Scheme
|
Habitat |
Season |
Suitability |
Major Importance? |
|
1.5. Forest -> Forest – Subtropical/Tropical Dry |
– |
Suitable |
– |
|
2.1. Savanna -> Savanna – Dry |
– |
Suitable |
– |
|
2.2. Savanna -> Savanna – Moist |
– |
Suitable |
– |
|
14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land |
– |
Suitable |
– |
|
14.2. Artificial/Terrestrial -> Artificial/Terrestrial – Pastureland |
– |
Suitable |
– |
|
14.3. Artificial/Terrestrial -> Artificial/Terrestrial – Plantations |
– |
Suitable |
– |
Life History
Generation Length: (Not specified)
Age at maturity: female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species may opportunistically be used as bushmeat (for example, in Zimbabwe; D. MacFadyen pers. obs. 2005), but this is not expected to impact the population.
|
Subsistence: |
Rationale: |
Local Commercial: |
Further detail including information on economic value if available: |
|
Yes |
– |
– |
– |
National Commercial Value: No
International Commercial Value: No
|
End Use |
Subsistence |
National |
International |
Other (please specify) |
|
1. Food – human |
true |
– |
– |
– |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
There are no major threats to this species, but it requires substantial cover so overgrazing, overstocking and incorrect fire regimes may negatively affect local population densities (Bowland and Perrin 1988, 1989).
Current habitat trend: Stable. Habitat will not decline as savannah habitat is well-protected within the assessment region and it can exist in agricultural landscapes. However, overstocking of livestock farms and wildlife ranches will affect habitat quality.
Conservation
It occurs in a number of protected areas, including Kruger National Park and Venetia Limpopo Nature Reserve. No specific interventions are necessary at present. However, protection of patches of natural habitat through biodiversity stewardship programmes and reduction in stocking rates to retain ground cover will benefit this species.
Recommendations for land managers and practitioners:
- Landowners and communities should be incentivised to stock livestock or wildlife at ecological carrying capacity and to implement a natural fire regime.
Research priorities:
- Its precise distributional limits are unknown and further sampling and molecular analysis are required to distinguish between the ranges of the two distinct Aethomys chrysophilus populations. This includes vetting of existing museum records.
- Ecological and behavioural interactions between these populations should be researched.
Encouraged citizen actions:
- Practise indigenous gardening to sustain small mammal diversity, especially in rural areas.
- Encourage corridors of indigenous, rank vegetation to connect areas of suitable habitat.
Bibliography
Avenant NL, Kuyler P. 2002. Small mammal diversity in the Maguga Dam inundation area, Swaziland. South African Journal of Wildlife Research 32: 101-108.
Bowland AE, Perrin MR. 1988. The effect of fire on the small mammal community in Hluhluwe Game Reserve. Mammalian Biology 53: 235–244.
Bowland AE, Perrin MR. 1989. The effect of overgrazing on the small mammals in Umfolozi Game Reserve. Mammalian Biology 54: 251–260.
Castiglia R, Corti M, Colangelo P, Annesi F, Capanna E, Verheyen W, Sichilima A, Makundi R. 2003. Chromosomal and molecular characterization of Aethomys kaiseri from Zambia and Aethomys chrysophilus from Tanzania (Rodentia, Muridae). Hereditas 139: 81–89.
Chimimba CT, Dippenaar NJ, Robinson TJ. 1999. Morphometric and morphological delineation of southern African species of Aethomys (Rodentia: Muridae). Biological Journal of the Linnean Society 67: 501–527.
Davis DH. 1962. Distribution patterns of southern African Muridae, with notes on some of their fossil antecedents. Annals of the Cape Province Museum of Natural History 2: 56–76.
Ducroz J-F, Volobouev V., Granjon L. 2001. An assessment of the systematics of Arcicanthine rodents using mitochondrial DNA sequences: evolutionary and biogeographical implications. Journal of Mammalian Evolution, 8, 173-206.
Ellerman JR 1941. The families and genera of living rodents. British Museum (Natural History), London.
Fuller JA, Perrin MR. 2001. Habitat assessment of small mammals in the Umvoti Vlei Conservancy, KwaZulu-Natal, South Africa. South African Journal of Wildlife Research 31: 1–12.
Lecompte E., Aplin, K., Denys, C., Catzeflis, F., Chades M., Chevret, P. 2008. Phylogeny and biogeography of African Murinae based on mitochondrial dan nuclear gene sequences, with a new tribal classification of the subfamily. BMC Evolutionary Biology, 8:199.
Linzey AV, Chimimba CT. 2008. Aethomys chrysophilus (Rodentia: Muridae). Mammalian Species 808: 1–10.
Linzey AV and Kesner MH. 1997a. Small mammals of a woodland-savannah ecosystem in Zimbabwe. I. Density and habitat occupancy patterns. Journal of Zoology 243(1): 137-152. https://doi.org/10.1111/j.14697998.1997.tb05760.x
Linzey AV and Kesner MH. 1997b. Small mammals of a woodland-savannah ecosystem in Zimbabwe. II. Community structure. Journal of Zoology 243(1): 153-162. https://doi.org/10.1111/j.1469-7998.1997.tb05761.x
Linzey AV, Kesner MH, Chimimba CT. 2013. Aethomys chrysophilus Red Veld Rat (Red Aethomys). Pages 365–366 in Happold DCD, editor. Mammals of Africa. Volume III: Rodents, Hares and Rabbits. Bloomsbury Publishing, London, UK.
MacFadyen D. 2007. A comparative study of rodent and shrew diversity and abundance in and outside the N’washitshumbe enclosure site in the Kruger National Park. M.Sc. Thesis. University of Pretoria.
Mamba, M.L., Da Conceicao, A.G., Naskrecki, P., Ngovene, A., Dalton, D.L., Russo, I.R.M., Visser, F. and Monadjem, A., 2024. An annotated checklist of the mammals of the Chimanimani National Park, Mozambique. 20 (5): 1222–1258. https://doi.org/10.15560/20.5.1222
Mazoch, V., Mikula, O., Bryja, J., Konvičková, H., Russo, I.R., Verheyen, E. and Šumbera, R., 2018. Phylogeography of a widespread sub-Saharan murid rodent Aethomys chrysophilus: the role of geographic barriers and paleoclimate in the Zambezian bioregion. Mammalia, 82(4), pp.373-387.
Monadjem, A., Taylor, P.J., Denys, C. and Cotterill, F.P.D. 2015. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis. De Gruyter, Berlin, Germany.
Musser GG, Carleton, MD (1993) Family Muridae. In: Mammal species of the world: a taxonomic and geographic reference, Second edition (eds. Wilson DE and Reeder DM), pp. 501-755. Smithsonian Institution Press, in association with the American Society of Mammalogists, London and Washington D.C.
Power RJ. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng, South Africa.
Skinner J.D. and Chimimba C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.
Wilson, DE., Lacher, TE., Mittermeier, RA. (eds). 2017. Handbook of the mammals of the world. Vol 7. Rodents II. Lynx Editions, Barcelona.