

Pygmy Sperm Whale
Kogia breviceps

2025 Red list status
Data Deficient
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Kogia breviceps – (Blainville, 1838)
ANIMALIA – CHORDATA – MAMMALIA – ARTIODACTYLA – KOGIIDAE – Kogia – breviceps
Common Names: Pygmy Sperm Whale, Lesser Cachalot, Lesser Sperm Whale (English), Dwergpotvis (Afrikaans), Cachalot pygmée (French), Cachalote cabeza chica (Spanish; Castilian), Cachalote pigmeo (Spanish; Castilian)
Synonyms: Kogia breviceps Blainville, 1838
Taxonomic Note:
The genus Kogia contains two species, the Pygmy Sperm Whale (Kogia breviceps) and Dwarf Sperm Whale (Kogia sima), with no subspecies recognised (Committee on Taxonomy 2021). The cryptic behavior and similar external morphology of the two species mean that records are often not displayed at the species level and are instead combined into Kogia spp. It has been suggested that K. sima may consist of two parapatric species occupying the Atlantic and Indo-Pacific Oceans, based on evidence from the mitochondrial cytochrome b gene (Chivers et al. 2005). K. breviceps and K. sima are clearly distinguished genetically, but full recognition of a putative third Kogia spp. awaits further supporting evidence (Plön et al. 2023: Chivers et al. 2005).
Red List Status: DD– Data Deficient
Assessment Information
Assessor: Plön, S.1
Reviewer: Patel, T.2
Institutions: 1 University of Cape Town, 2Endangered Wildlife Trust
Previous Assessors & Reviewers: Plön, S. & Relton, C.
Previous Contributors: Elwen, S., Findlay, K., Meÿer, M., Oosthuizen, H., Child, MF. & Taylor et al. (2008a, 2008b)
Assessment Rationale
This species is naturally uncommon and there are no regional estimates of population size or trends (Plön, 2022). However, as it is a deep-diving species (up to 800 m), the effects of marine noise pollution should be monitored (although thus far no strandings have been linked to this threat), and we urge more research into the severity of this threat within South African waters. Additionally, plastic pollution should be recognised as an increasing emerging threat, as they are known to fatally ingest plastic bags mistaken for squid. While their offshore distributions do not overlap with many major threats in the assessment region, their natural scarcity and presumed low population numbers make them vulnerable to minor threats, which may be increasing in severity in the assessment region. Thus, in line with the global assessments, we list this species as Data Deficient and urge more systematic monitoring and analysis to determine population sizes and trends within the assessment region.
Regional population effects: Pygmy Sperm Whales occur extensively in pelagic waters of tropical and temperate regions, and although their movement patterns are largely unknown, no barriers to dispersal have been recognised (Plön et al. 2023), thus rescue effects are considered possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Plön S. 2025. A conservation assessment of Kogia breviceps. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
Both Pygmy Sperm Whales and Dwarf Sperm Whales are widely distributed and inhabit deep tropical, subtropical and temperate waters throughout all oceans (Plön, 2022; Plön and Baird, 2022; McAlpine 2002). However, the Pygmy Sperm Whale seems to occur more commonly in cooler temperate regions in comparison to the Dwarf Sperm Whale, which seemingly prefers warmer waters (Plön and Baird, 2022; Caldwell & Caldwell 1989). The distributional ranges of Kogia spp. are poorly known, and most records originate from strandings or occasionally as individuals captured in small fisheries, rather than live sightings at sea (Plön, 2022; Nagorsen 1985; Caldwell & Caldwell 1989; McAlpine 2002). This may, however, be attributed to their cryptic nature and affiliation for pelagic regions, rather than low population abundances, particularly since a number of recent studies report extensive observations of K. breviceps in the wild off Hawaii (Baird, 2005) and off the Canary Islands (Vidal et al. 2024). The Pygmy Sperm Whale is thought to predominantly reside in pelagic regions, beyond the continental shelf edge (Plön, 2022; Ross 1979).
Within the assessment region, stranding records of Pygmy Sperm Whales suggest that this species occurs from 22°S on South Africa’s west coast (18°33’ S on the west coast of Southern Africa (Elwen et al. 2013) to 33°42’S on the east coast (Plön et al. 2023; Findlay et al. 1992). Prior to 1977, Ross (1979) documented strandings of 42 Dwarf Sperm Whales along South Africa’s coastline from Saldanha Bay to East London. These and additional records suggest that this species is restricted to South Africa’s southern coast between 17.8°E and 28°E (Ross 1979; Findlay et al. 1992), and an outlying record from KwaZulu-Natal is thought to be attributed to unusual environmental conditions (Ross et al. 1985). The lack of records northwards from Saldanha Bay and along the east coast of South Africa suggests that Dwarf Sperm Whales may prefer the mixed-water conditions, where the Benguela and Agulhas Currents mix (Ross 1984). Due to the rarity of sightings at sea, distribution in the subregion has to be largely deduced from the incidence of strandings (Best, 2007), which have occurred throughout the year in the assessment region.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
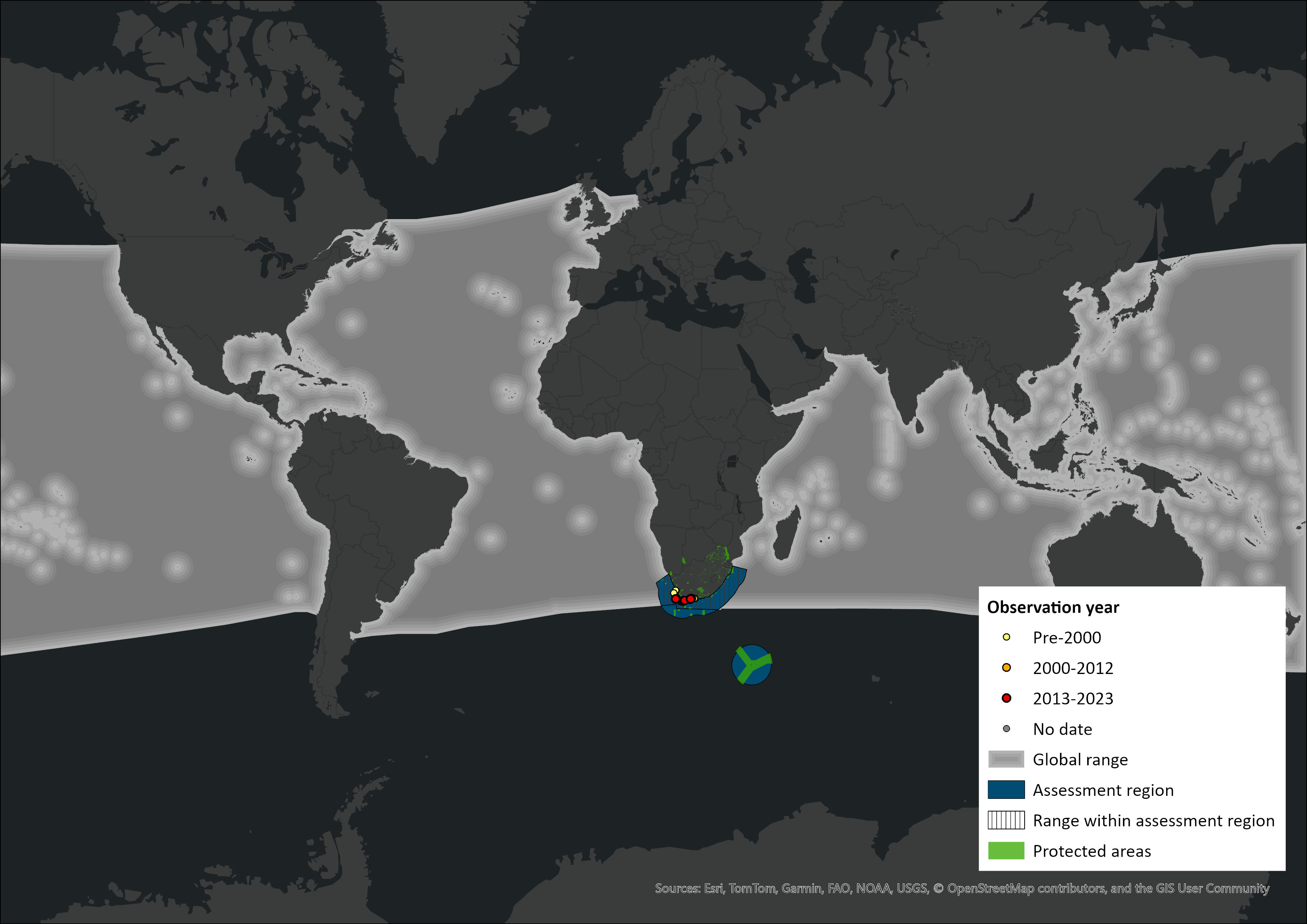 Figure 1. Distribution records for Pygmy Sperm Whale (Kogia breviceps) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Pygmy Sperm Whale (Kogia breviceps) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical, Australasian, Indomalayan, Nearctic, Neotropical, Oceanian, Palearctic
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| American Samoa | Extant | Native | – | Resident |
| Angola | Extant | Native | – | Resident |
| Anguilla | Extant | Native | – | Resident |
| Antigua and Barbuda | Extant | Native | – | Resident |
| Argentina | Extant | Native | – | Resident |
| Aruba | Extant | Native | – | Resident |
| Australia | Extant | Native | – | Resident |
| Australia -> Tasmania | Extant | Native | – | Resident |
| Bahamas | Extant | Native | – | Resident |
| Bangladesh | Extant | Native | – | Resident |
| Barbados | Extant | Native | – | Resident |
| Belgium | Extant | Native | – | Resident |
| Belize | Extant | Native | – | Resident |
| Benin | Extant | Native | – | Resident |
| Bermuda | Extant | Native | – | Resident |
| Bonaire, Sint Eustatius and Saba | Extant | Native | – | Resident |
| Brazil | Extant | Native | – | Resident |
| British Indian Ocean Territory | Extant | Native | – | Resident |
| Brunei Darussalam | Extant | Native | – | Resident |
| Cabo Verde | Extant | Native | – | Resident |
| Cambodia | Extant | Native | – | Resident |
| Cameroon | Extant | Native | – | Resident |
| Canada | Extant | Native | – | Resident |
| Canada -> Nova Scotia | Extant | Native | – | Resident |
| Cayman Islands | Extant | Native | – | Resident |
| Chile | Extant | Native | – | Resident |
| Chile -> Juan Fernández Is. | Extant | Native | – | Resident |
| China | Extant | Native | – | Resident |
| Christmas Island | Extant | Native | – | Resident |
| Cocos (Keeling) Islands | Extant | Native | – | Resident |
| Colombia | Extant | Native | – | Resident |
| Comoros | Extant | Native | – | Resident |
| Congo | Extant | Native | – | Resident |
| Congo, The Democratic Republic of the | Extant | Native | – | Resident |
| Cook Islands | Extant | Native | – | Resident |
| Costa Rica | Extant | Native | – | Resident |
| Cuba | Extant | Native | – | Resident |
| Curaçao | Extant | Native | – | Resident |
| Côte d’Ivoire | Extant | Native | – | Resident |
| Denmark | Extant | Native | – | Resident |
| Djibouti | Extant | Native | – | Resident |
| Dominica | Extant | Native | – | Resident |
| Dominican Republic | Extant | Native | – | Resident |
| Ecuador | Extant | Native | – | Resident |
| El Salvador | Extant | Native | – | Resident |
| Equatorial Guinea | Extant | Native | – | Resident |
| Eritrea | Presence Uncertain | Native | – | Seasonal Occurrence Uncertain |
| Fiji | Extant | Native | – | Resident |
| France | Extant | Native | – | Resident |
| French Guiana | Extant | Native | – | Resident |
| French Polynesia | Extant | Native | – | Resident |
| Gabon | Extant | Native | – | Resident |
| Gambia | Extant | Native | – | Resident |
| Germany | Extant | Native | – | Resident |
| Ghana | Extant | Native | – | Resident |
| Gibraltar | Extant | Native | – | Resident |
| Grenada | Extant | Native | – | Resident |
| Guadeloupe | Extant | Native | – | Resident |
| Guam | Extant | Native | – | Resident |
| Guatemala | Extant | Native | – | Resident |
| Guernsey | Extant | Native | – | Resident |
| Guinea | Extant | Native | – | Resident |
| Guinea-Bissau | Extant | Native | – | Resident |
| Guyana | Extant | Native | – | Resident |
| Haiti | Extant | Native | – | Resident |
| Honduras | Extant | Native | – | Resident |
| Hong Kong | Extant | Native | – | Resident |
| India | Extant | Native | – | Resident |
| Indonesia | Extant | Native | – | Resident |
| Iran, Islamic Republic of | Extant | Native | – | Resident |
| Ireland | Extant | Native | – | Resident |
| Isle of Man | Extant | Native | – | Resident |
| Jamaica | Extant | Native | – | Resident |
| Japan | Extant | Native | – | Resident |
| Japan -> Honshu | Extant | Native | – | Resident |
| Jersey | Extant | Native | – | Resident |
| Kenya | Extant | Native | – | Resident |
| Kiribati | Extant | Native | – | Resident |
| Korea, Democratic People’s Republic of | Extant | Native | – | Resident |
| Korea, Republic of | Extant | Native | – | Resident |
| Liberia | Extant | Native | – | Resident |
| Macao | Extant | Native | – | Resident |
| Madagascar | Extant | Native | – | Resident |
| Malaysia | Extant | Native | – | Resident |
| Maldives | Extant | Native | – | Resident |
| Marshall Islands | Extant | Native | – | Resident |
| Martinique | Extant | Native | – | Resident |
| Mauritania | Extant | Native | – | Resident |
| Mauritius | Extant | Native | – | Resident |
| Mayotte | Extant | Native | – | Resident |
| Mexico | Extant | Native | – | Resident |
| Micronesia, Federated States of | Extant | Native | – | Resident |
| Montserrat | Extant | Native | – | Resident |
| Morocco | Extant | Native | – | Resident |
| Mozambique | Extant | Native | – | Resident |
| Myanmar | Extant | Native | – | Resident |
| Namibia | Extant | Native | – | Resident |
| Nauru | Extant | Native | – | Resident |
| Netherlands | Extant | Native | – | Resident |
| New Caledonia | Extant | Native | – | Resident |
| New Zealand | Extant | Native | – | Resident |
| Nicaragua | Extant | Native | – | Resident |
| Nigeria | Extant | Native | – | Resident |
| Niue | Extant | Native | – | Resident |
| Norfolk Island | Extant | Native | – | Resident |
| Northern Mariana Islands | Extant | Native | – | Resident |
| Oman | Extant | Native | – | Resident |
| Pakistan | Extant | Native | – | Resident |
| Palau | Extant | Native | – | Resident |
| Panama | Extant | Native | – | Resident |
| Papua New Guinea | Extant | Native | – | Resident |
| Peru | Extant | Native | – | Resident |
| Philippines | Extant | Native | – | Resident |
| Pitcairn | Extant | Native | – | Resident |
| Portugal | Extant | Native | – | Resident |
| Portugal -> Azores | Extant | Native | – | Resident |
| Puerto Rico | Extant | Native | – | Resident |
| Russian Federation | Presence Uncertain | Native | – | Seasonal Occurrence Uncertain |
| Réunion | Extant | Native | – | Resident |
| Saint Barthélemy | Extant | Native | – | Resident |
| Saint Helena, Ascension and Tristan da Cunha | Extant | Native | – | Resident |
| Saint Kitts and Nevis | Extant | Native | – | Resident |
| Saint Lucia | Extant | Native | – | Resident |
| Saint Martin (French part) | Extant | Native | – | Resident |
| Saint Pierre and Miquelon | Extant | Native | – | Resident |
| Saint Vincent and the Grenadines | Extant | Native | – | Resident |
| Samoa | Extant | Native | – | Resident |
| Sao Tome and Principe | Extant | Native | – | Resident |
| Senegal | Extant | Native | – | Resident |
| Seychelles | Extant | Native | – | Resident |
| Sierra Leone | Extant | Native | – | Resident |
| Singapore | Extant | Native | – | Resident |
| Sint Maarten (Dutch part) | Extant | Native | – | Resident |
| Solomon Islands | Extant | Native | – | Resident |
| Somalia | Extant | Native | – | Resident |
| South Africa | Extant | Native | – | Resident |
| South Africa -> Eastern Cape Province | Extant | Native | – | Resident |
| South Africa -> KwaZulu-Natal | Extant | Native | – | Resident |
| South Africa -> Northern Cape Province | Extant | Native | – | Resident |
| South Africa -> Western Cape | Extant | Native | – | Resident |
| Spain | Extant | Native | – | Resident |
| Sri Lanka | Extant | Native | – | Resident |
| Suriname | Extant | Native | – | Resident |
| Taiwan, Province of China | Extant | Native | – | Resident |
| Tanzania, United Republic of | Extant | Native | – | Resident |
| Thailand | Extant | Native | – | Resident |
| Timor-Leste | Extant | Native | – | Resident |
| Togo | Extant | Native | – | Resident |
| Tokelau | Extant | Native | – | Resident |
| Tonga | Extant | Native | – | Resident |
| Trinidad and Tobago | Extant | Native | – | Resident |
| Turks and Caicos Islands | Extant | Native | – | Resident |
| Tuvalu | Extant | Native | – | Resident |
| United Arab Emirates | Extant | Native | – | Resident |
| United Kingdom of Great Britain and Northern Ireland | Extant | Native | – | Resident |
| United States of America | Extant | Native | – | Resident |
| United States of America -> Hawaiian Is. | Extant | Native | – | Resident |
| United States of America -> Washington | Extant | Native | – | Resident |
| Uruguay | Extant | Native | – | Resident |
| Vanuatu | Extant | Native | – | Resident |
| Venezuela, Bolivarian Republic of | Extant | Native | – | Resident |
| Viet Nam | Extant | Native | – | Resident |
| Virgin Islands, British | Extant | Native | – | Resident |
| Virgin Islands, U.S. | Extant | Native | – | Resident |
| Wallis and Futuna | Extant | Native | – | Resident |
| Western Sahara | Extant | Native | – | Resident |
| Yemen | Extant | Native | – | Resident |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
| Presence | Origin | Formerly Bred | Seasonality | |
| 21. Atlantic – northwest | Extant | Native | – | – |
| 27. Atlantic – northeast | Extant | Native | – | – |
| 31. Atlantic – western central | Extant | Native | – | – |
| 34. Atlantic – eastern central | Extant | Native | – | – |
| 41. Atlantic – southwest | Extant | Native | – | – |
| 47. Atlantic – southeast | Extant | Native | – | – |
| 51. Indian Ocean – western | Extant | Native | – | – |
| 57. Indian Ocean – eastern | Extant | Native | – | – |
| 61. Pacific – northwest | Extant | Native | – | – |
| 67. Pacific – northeast | Extant | Native | – | – |
| 71. Pacific – western central | Extant | Native | – | – |
| 77. Pacific – eastern central | Extant | Native | – | – |
| 81. Pacific – southwest | Extant | Native | – | – |
| 87. Pacific – southeast | Extant | Native | – | – |
Climate change
The impact of global climate change, and the associated effects of increased water temperature and CO2 concentration on Kogia spp. is largely unknown; however, it is likely to have cascading effects on the movements and feeding ecology of these species (Learmonth et al. 2006).
Population information
Abundance estimates of this species is often underestimated due to their offshore habitats, long and deep-diving behaviour and inconspicuous nature at the surface (Barlow 1999). Additionally, Pygmy and Dwarf Sperm Whales are often confused during sightings, which further complicates any population assessments. Although no global population abundance estimates are available for the species, the sum of the available abundance estimates from various geographical regions is over 10,000 individuals, but because the surveys only covered a small fraction of the entire habitat, the total global abundance is likely much higher (Kiszka and Braulik 2020).
However, the stranding frequency of K. breviceps in regions such as South Africa and the southeastern United States suggests they may be more common than sighting records would imply (Plön, 2022; Hodge et al. 2018). Similarly, although no regional population estimates exist for K. breviceps in the southern hemisphere, there appears to be substantial gene flow between stocks (Plön et al. 2023). The species is primarily known from stranding records, which are declining across the region. However, because the cause of strandings is largely unknown, it is unclear how this trend is reflective of the population.
The 3-generation period of the species is calculated as 36 years (Taylor et al. 2007), and, globally, a 30% reduction over three generations cannot be ruled out (Taylor et al. 2008a, 2008b).
Current population trend: Unknown
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Unknown
Severely fragmented: No
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
Molecular evidence revealed extensive genetic mixing across the global distribution of this species indicating it is a single metapopulation (Plön et al., 2023). Given that no estimates of population size are available and no measure of effective population size (Ne) was conducted using the available genetic information, it is not possible to quantify the Ne 500 indicator.
Habitats and ecology
Pygmy Sperm Whales inhabit deep waters over the continental shelf and slope (Plön, 2022; Ross 1979). Although some studies document some division between K. breviceps and K. sima in their preference for water depth, largely based on the analysis of stomach contents (Plön, 2004; Ross, 1979), Mullin et al. (1994) using aerial observations noted that in the Gulf of Mexico, both species occurred in waters between 400–600 m deep. These depths constituted the upper regions of the continental slope, which exhibited high concentrations of zooplankton (Baumgartner et al. 2001). Both species appear to feed over the deeper continental shelf and slope (Plön, 2022; Ross 1979).
Ross (1979) reported that Pygmy Sperm Whales may occur individually or in small groups of up to six animals in the assessment region. Although they are capable of rapid bursts in speed, they typically swim at about 5.5 km / h (Mörzer Bruyns 1971). While not commonly sighted at sea, Katona et al. (1983) reported that the species is easy to approach, as they often float on the surface of the water with much of their body exposed (Leatherwood & Reeves 1983).
The stomach contents of stranded Pygmy Sperm Whales contained the remains of deep-water cephalopods, suggesting that this species feeds at depths of up to 900 m (Plön, 2022). The majority of the diet of Kogia spp. consists of cephalopods, but they may also consume other prey, for example deep-sea fishes and shrimps (McAlpine et al. 1997; dos Santos & Haimovici 2001; S. Plön pers. comm.). In South African waters, Pygmy and Dwarf Sperm Whales consumed at least 67 and 38 different prey species, respectively, and Pygmy Sperm Whales are likely to feed in deeper waters compared to Dwarf Sperm Whales (Plön 2004; Ross 1979).
Although little is known about the reproductive biology of Kogia spp., and no seasonality can be inferred, female and male K. breviceps from South Africa are estimated to reach sexual maturity at 2.6 m and 2.4 m, respectively (Plön, 2004). Calves are born at an approximate length of 1.2 m (Plön, 2004). Surprisingly, an extremely high proportion (80%) of calves and foetuses were found to be male (Ross 1979).
Ecosystem and cultural services: Marine mammals integrate and reflect ecological variation across large spatial and long temporal scales, and therefore they are prime sentinels of marine ecosystem change (Moore 2008).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 10.1. Marine Oceanic -> Marine Oceanic – Epipelagic (0-200m) | – | Suitable | Yes |
| 10.2. Marine Oceanic -> Marine Oceanic – Mesopelagic (200-1000m) | – | Suitable | Yes |
| 10.3. Marine Oceanic -> Marine Oceanic – Bathypelagic (1000-4000m) | – | Suitable | Yes |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: 5 yrs (Plön, 2004)
Age at Maturity: Male: 2.5-5 yrs (Plön, 2004)
Size at Maturity (in cms): Female: 262 cm (Plön, 2004)
Size at Maturity (in cms): Male: 241-242 cm (Plön, 2004)
Longevity: 23 yrs in females, 16 yrs in males (Plön, 2004)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): 324cm in females, 275cm in males (Plön, 2004)
Size at Birth (in cms): 120cm (Plön, 2004)
Gestation Time: 11-12 months (Plön, 2004)
Reproductive Periodicity: Annual reproduction (Plön, 2004)
Average Annual Fecundity or Litter Size: 1
Natural Mortality: (Not specified)
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Marine
General Use and Trade Information
Although this species is hunted at low levels in some parts of their range, there is no trade or use of the species within the assessment region. In other parts of the world, such as the Philippines, Kogia spp. are hunted for bait to be used in fisheries or meat for human consumption (Leatherwood et al. 1992; Anonymous 1996). Additionally, where they are caught accidentally in fisheries, such as gillnets, they may also be utilised for human consumption (Klinowska 1991; Muñoz-Hincapié et al. 1998).
|
Subsistence: |
Rationale: |
Local Commercial: |
Further detail including information on economic value if available: |
|
Yes |
– |
– |
– |
National Commercial Value: Yes
International Commercial Value: No
|
End Use |
Subsistence |
National |
International |
Other (please specify) |
|
1. Food – human |
true |
true |
– |
– |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
Although no major threats have been recognised for Kogia spp. within the assessment region, and the threats listed below are not likely to cause drastic population reduction on their own, they may result in slow, significant declines in the future, especially if the threats synergise.
Kogia spp. were not historically hunted commercially, but are currently hunted on a small-scale in regions such as Japan, Taiwan, Sri Lanka, Indonesia and the Lesser Antilles (Jefferson et al. 1993), and there are reports of sporadic accidental bycatch of Pygmy Sperm Whales in gillnet, longline and purse seine fisheries across the Northern Hemisphere (Jefferson et al. 1993; Barlow et al. 1997; Perez et al. 2001; Garrison 2007). No direct or indirect catches of Kogia species have been reported from South African waters, and Baird et al. (1996) suggests that direct and indirect catches of the Pygmy Sperm Whale are not expected to severely impact their population stability.
The ingestion of plastic bags (which the animals possibly mistake for squid) is common among squid-eating cetaceans and has been documented for both Kogia species (for example, Stamper et al. 2006), and is considered to be relatively common among these species (Plön 2022; Plön & Baird 2022). Plastic pollution in the stomachs of cetaceans frequently hinders natural digestion of food resources, leading to gut-blockage, starvation, strandings and death (Caldwell & Caldwell 1989; Laist et al. 1999; S. Plön pers. obs.).
As deep-diving species (similar to beaked whales), Pygmy and Dwarf Sperm Whales are presumably vulnerable to anthropogenic noise pollution, for example those produced by seismic surveys and sonar generated during naval operations (Cox et al. 2006; Wang & Yang 2006; Yang et al. 2008). A number of stranding events, which included Kogia spp., have been documented in Taiwan (Wang & Yang 2006; Yang et al. 2008), the Gulf of Mexico and off the east coast of Florida (Waring et al. 2006). Although, anthropogenic noise pollution is a possible cause of these stranding incidents, due to spatial and temporal associations, this assumption has not been confirmed (Hohn et al. 2006; Wang & Yang 2006; Yang et al. 2008). Marine noise pollution is thought to be intensifying within South African waters (Koper & Plön 2012).
Conservation
K. brevicepsis listed in Appendix II of the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES). In the absence of further data, no specific conservation measures can be recommended for this species within the assessment region at present. Continued research is necessary into the impact of and interactions between threats, such as plastic and noise pollution, climate change and incidental bycatch in pelagic fisheries. Management plans should be developed as such data become avail.
Recommendations for managers and practitioners:
- The interaction between Kogia spp. and gillnet, purse seine and longline fisheries requires ongoing monitoring, and regular records of bycatch should be collected by fishing operations.
Research priorities:
- Population size, distribution and trends of these species in South African waters.
- Investigations into threats to these species, such as the impacts and severity of marine pollution (both plastics and noise) on populations.
Encouraged citizen actions:
- Use information dispensed by the South African Sustainable Seafood Initiative to make good choices when buying fish in shops and restaurants, e.g. wwfsa.mobi, FishMS 0794998795.
- Save electricity and fuel to mitigate CO2 emissions and hence, the rate of climate change.
- Sightings data from pelagic commercial tourism operators may be valuable.
- Report any strandings to the relevant local authorities.
Bibliography
Baird, R. W. (2005). Sightings of dwarf (Kogia sima) and pygmy (K. breviceps) sperm whales from the main Hawaiian Islands. Pacific Science, 59, 461-466.
Baird, R. W., Nelson, D., Lien, J. and Nagorsen, D. W. 1996. The status of the pygmy sperm whale, Kogia breviceps, in Canada. Canadian Field-Naturalist 110: 525-532.
Barlow, J. 1999. Trackline detection probability for long-diving whales. In: G. W. Garner, S. C. Amstrup, J. L. Laake, B. J. F. Manley, L. L. McDonald and D. G. Robertson (eds), Marine mammal survey and assessment methods, pp. 209-221. Balkema Press, Netherlands.
Barlow, J., Forney, K. A., Hill, K. A., Brownell Jr., R. L., Caretta, R. L., Demaster, D. P., Julian, D. P., Lowry, M. S., Ragen, M. S. and Reeves, R. R. 1997. U.S. Pacific marine mammal stock assessments: 1996. NOAA Technical Memorandum NMFS-SWFSC 248: 223 pp.
Baumgartner, M. F., Mullin, K. D., May, L. N. and Leming, T. D. 2001. Cetacean habitats in the northern Gulf of Mexico. Fishery Bulletin 99: 219-239.
Best, P. B. B. 2007. Whales and Dolphins of the Southern African Subregion. Cambridge University Press, Cape Town. 338pp.
Caldwell, D. K. and Caldwell, M. C. 1989. Pygmy sperm whale Kogia breviceps (de Blainville, 1838): Dwarf sperm whale Kogia simus Owen, 1866. In: S. H. Ridgway and R. Harrison (eds), Handbook of marine mammals, Vol. 4: River dolphins and the larger toothed whales, pp. 234-260. Academic Press.
Chivers, S.J., Leduc, A.E., Robertson, K.M., Barros, N.B. and Dizon, A.E. 2005. Genetic variation in Kogia spp., with preliminary evidence for two species of Kogia sima. Marine Mammal Science 21(4): 619-634.
Cox, T.M., Ragen, T.J., Read, A.J., Vos, E., Baird, R.W., Balcomb, K., Barlow, J., Caldwell, J., Cranford, T., Crum, L., D’Amico, A., D’Spain, A., Fernández, J., Finneran, J., Gentry, R., Gerth, W., Gulland, F., Hildebrand, J., Houser, D., Hullar, T., Jepson, P.D., Ketten, D., Macleod, C.D., Miller, P., Moore, S., Mountain, D., Palka, D., Ponganis, P., Rommel, S., Rowles, T., Taylor, B., Tyack, P., Wartzok, D., Gisiner, R., Mead, J. and Benner, L. 2006. Understanding the impacts of anthropogenic sound on beaked whales. Journal of Cetacean Research and Management 7(3): 177-187.
Elwen, S.H., Gridley, T., Roux, J.-P., Best P. B. and Smale, M. J. 2013. Records of kogiid whales in Namibia, including the first record of the dwarf sperm whale (Kogia sima). Marine Biodiversity Records 6: e45.
Findlay KP, Best PB, Ross GJB, Cockcroft VG. 1992. The distribution of small odontocete cetaceans off the coasts of South Africa and Namibia. South African Journal of Marine Science 12: 237-270.
Garrison LP. 2007. Interactions between marine mammals and pelagic longline fishing gear in the US Atlantic Ocean between 1992 and 2004. Fishery Bulletin 105: 408–417.
Handley, C.O. 1966. A synopsis of the genus Kogia (pygmy sperm whales). In: K.S. Norris (ed.), Whales, dolphins, and porpoises, pp. 62-69. University of California Press, Berkeley, California, USA.
Hodge, L. E. W., Baumann-Pickering, S., Hildebrand, J. A., Bell, J. T., Cummings, E. W., Foley, H. J., McAlarney, R. J., McLellan, W. A., Pabst, D. A., Swaim, Z. T., Waples, D. M., & Read, A. J. (2018). Heard but not seen: Occurrence of Kogia spp. along the western North Atlantic shelf break. Marine Mammal Science 34: 1141-1153.
Hohn, A.A., Rotstein, D.S., Harms, C.A. and Southall, B.L. 2006. Multispecies mass stranding of pilot whales (Globicephala macrorhynchus), minke whale (Balaenoptera acutorostrata), and dwarf sperm whales (Kogia sima) in North Carolina on 15-16 January 2005. NOAA Technical Memorandum NMFS-SEFSC-57.
IUCN (International Union for Conservation of Nature). 2012a. Kogia breviceps. Available at: http://www.iucnredlist.org. (Accessed: 21 February 2016).
IUCN (International Union for Conservation of Nature). 2012b.. Kogia sima. Available at: http://www.iucnredlist.org. (Accessed: 21 February 2016).
Jefferson, T. A., Leatherwood, S. and Webber, M. A. 1993. Marine Mammals of the World: FAO Species Identification Guide. United Nation Environment Programme and Food and Agricultural Organization of the UN.
Katona SK, Rough V, Richardson DT. 1983. A field guide to the whales, porpoises and seals of the Gulf of Maine and Eastern Canada, Cape Cod to Newfoundland. Charles Scribners Sons, New York, USA.
Kiszka J, Braulik G (2020) Kogia breviceps. The IUCN Red List of Threatened Species 2020: e. T11047A50358334. https://doi.org/10.2305/IUCN. UK.2020-2.RLTS.T11047A50358334.en
Klinowska, M. 1991. Dolphins, Porpoises and Whales of the World. The IUCN Red Data Book. IUCN, Gland and Cambridge. viii + 429pp.
Koper RP, Plön S. 2012. The potential impacts of anthropogenic noise on marine animals and recommendations for research in South Africa. EWT Research & Technical Paper No. 1. Endangered Wildlife Trust, South Africa.
Laist, D. W., Coe, J. M. and O’Hara, K. J. 1999. Marine debris pollution. In: J. R. Twiss and R. R. Reeves (eds), Conservation and Management of Marine Mammals.
Learmonth, J.A., Macleod, C.D., Santos, M.B., Pierce, G.J., Crick, H.Q.P. and Robinson, R.A. 2006. Potential effects of climate change on marine mammals. Oceanography and Marine Biology: An Annual Review 44: 431-464.
Leatherwood S, Dolar MLL, Wood CJ, Aragones LV, Hill CL. 1992. Marine mammal species confirmed from Philippine waters. Silliman Journal 36: 65–81.
Leatherwood S, Reeves RR. 1983. The Sierra Club Handbook of Whales and Dolphins. Sierra Club Books, San Francisco. USA.
Martin, V., Tejedor, M., Carrillo, M., Pérez-Gil, M., Arbelo, M., Servidio, A., Pérez-Gil, E.,Varo-Cruz, N., Fusar Poli, F., Aliart, S., Tejera, G., Lorente, M. and A. Fernández. (2024). Strandings and at sea observations reveal the canary archipelago as an important habitat for pygmy and dwarf sperm whale. Advances in Marine Biology, 99, 21-64.
McAlpine, D. F. 2002. Pygmy and dwarf sperm whales Kogia breviceps and K. simus. In: W. F. Perrin, B. Wursig and J. G. M. Thewissen (eds), Encyclopedia of Marine Mammals, pp. 1007-1009. Academic Press.
McAlpine, D. F., Murison, L. D. and Hoberg, E. P. 1997. New records for the pygmy sperm whale, Kogia breviceps (Physeteridae) from Atlantic Canada with notes on diet and parasites. Marine Mammal Science 13(4): 701-704.
Moore SE. 2008. Marine mammals as ecosystem sentinels. Journal of Mammalogy 89: 534-540.
Mullin, K. D., W. Hoggard, C. L. Roden, R. Lohoefener, C. M. Rogers, B. Taggart. 1994. Cetaceans on the upper continental slope in the north-central Gulf of Mexico. Fishery Bulletin 92: 773-786.
Muñoz-Hincapié MF, Mora-Pinto DM, Palacios DM, Secchi ER, Mignucci-Giannoni AA. 1998. First osteological record of the dwarf sperm whale in Colombia, with notes on the zoogeography of Kogia in South America. Revista de la Academia Colombiana de Ciencias 22: 433–44.
Mörzer Bruyns WFJ. 1971. Field Guide to Whales and Dolphins. Mees, Amsterdam.
Nagorsen, D. 1985. Kogia simus. Mammalian Species 239: 1-6.
Perez, C., Lopez, A., Sequeira, M., Silva, M., Herrera, R., Goncalves, J., Valdes, P., Mons, L., Freitag, L., Lens, S. and Cendero, O. 2001. Stranding and by-catch of cetaceans in the northeastern Atlantic during 1996.: 4pp. Copenhagen, Denmark.
Perrin, W. F., Dolar, M. L. L. and Alava, M. N. R. (eds). 1996. Report of the Workshop on the Biology and Conservation of Small Cetaceans and Dugongs of Southeast Asia. UNEP Report UNEP(W)/EAS WG, pp. 101 pp..
Plön S. 2004. The status and natural history of pygmy (Kogia breviceps) and dwarf (K. sima) sperm whales off Southern Africa. PhD thesis. Department of Zoology & Entomology, Rhodes University, South Africa. 553pp.
Plön, S., Best, P. B., Duignan, P., Lavery, S. D., Bernard, R. T. F., Van Waerebeek, K. and C. S. Baker. 2023. Population structure of pygmy (Kogia breviceps) and dwarf (Kogia sima) sperm whales in the Southern Hemisphere may reflect foraging ecology and dispersal patterns. Advances in Marine Biology 96, Vol. 1: 85-114.
Plön, S. 2022. Pygmy Sperm Whale Kogia breviceps (de Blainville, 1838). In: Hackländer, K., Zachos, F.E. (eds) Handbook of the Mammals of Europe. Handbook of the Mammals of Europe. Springer, Cham. doi.org/10.1007/978-3-319-65038-8_90-1.
Plön, S., Baird, R.W. 2022. Dwarf Sperm Whale, Kogia sima (Owen, 1866). In: Hackländer, K., Zachos, F.E. (eds) Handbook of the Mammals of Europe. Handbook of the Mammals of Europe. Springer, Cham. doi.org/10.1007/978-3-319-65038-8_91-1.
Rice, D.W. 1998. Marine Mammals of the World: Systematics and Distribution. Society for Marine Mammalogy, Special Publication Number 4, Lawrence, Kansas.
Ross GJ. 1984. Smaller cetaceans of the south east coast of southern Africa. Annals of the Cape Provincial Museums. Natural History 16: 309-319.
Ross GJB, Cockcroft VG, Cliff G. 1985. Additions to marine faunas of South Africa and Natal. Lammergeyer 35: 36–40.
Ross, G. J. B. 1979. Records of pygmy and dwarf sperm whales, genus Kogia, from southern Africa, with biological notes and some comparisons. Annals of the Cape Provincial Museums (Natural History) 11(14): 259-327.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Stamper, M. A., Whitaker, B. R., and Schofield, T. D. 2006. Case study: morbidity in a pygmy sperm whale Kogia breviceps due to ocean-borne plastic. Marine Mammal Science 22: 719-722.
Tarpley, R.J. and Marwitz, S. 1993. Plastic debris ingestion by cetaceans along the Texas coast: two case reports. Aquatic Mammals 19: 93–98.
Taylor, B. L., Baird, R., Barlow, J., Dawson, S. M., Ford, J., Mead, J. G., Notarbartolo di Sciara, G., Wade, P., and Pitman, R. L. 2008. Kogia sima. Available at: www.iucnredlist.org. (Accessed: 11 November 2008).
Taylor, B.L., Chivers, S.J., Larese, J. and Perrin, W.F. 2007. Generation length and percent mature estimates for IUCN assessments of cetaceans. NOAA, Southwest Fisheries Science Center Administrative Report LJ-07-01. La Jolla, California.
Wang, J.Y. and Yang, S.C. 2006. Unusual cetacean stranding events of Taiwan in 2004 and 2005. Journal of Cetacean Research and Management 8: 283-292.
Waring, G.T., Josephson, E., Fairfield, C.P. and Maze-Foley, K. 2006. U.S. Atlantic and Gulf of Mexico marine mammal stock assessments – 2005. NOAA Technical Memorandum NMFS-NE-201. 346 p.
Yamada M. 1954. An account of a rare porpoise, Feresa Gray from Japan. Scientific Reports of the Whales Research Institute, Tokyo 9: 59-88.
Yang, W.-C., Chou, L.-S., Jepson, P. D., Brownell Jr., R. L., Cowan, D., Chang, P.-H., Chiou, H.-I., Yao, C.-J., Yamada, T. K., Chiu, J.-T., Wang, P.-J. and Fernandez, A. 2008. Unusual cetacean mortality events in Taiwan, possibly linked to naval activities. Veterinary Record 162: 184-186.
Zerbini, A. N. and Kotas, J. E. 1998. A note on cetacean bycatch in pelagic driftnetting off southern Brazil. Reports of the International Whaling Commission 48: 519-524.
dos Santos, RA. and Haimovici, M. 2001. Cephalopods in the diet of marine mammals stranded or incidentally caught along southeastern and southern Brazil (21- 34º S). Fisheries Research 52: 99-112.