

Peak-saddle Horseshoe Bat
Rhinolophus blasii

2025 Red list status
Near threatened
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Rhinolophus blasii – Peters, 1867
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – RHINOLOPHIDAE – Rhinolophus – blasii
Common Names: Peak-saddle Horseshoe Bat, Blasius’s Horseshoe Bat, Peters’s Horseshoe Bat (English), Spitssaalneusvlermuis (Afrikaans)
Synonyms: No Synonyms
Taxonomic Note:
The taxonomic status of the putative subspecies, Rhinolophus blasii empusa (K. Anderson 1904) needs revision. The type specimen for the species is from Italy (ZMB 557, Syntype), while the type locality of the southern African R. b. empusa is Zomba, Malawi (BM 93.7.9.33, Holotype) (Monadjem et al. 2020). A phylogeographic study on the Rhinolophidae identified two clades within R. blasii in Africa, a northern clade including Iran, Algeria and Morrocco, and a southeastern clade compromised of individuals from Liberia, Tanzania, Malawi, Mozambique, Zimbabwe and South Africa (Demos et al. 2019).
Red List Status: NT – Near Threatened D1 (IUCN version 3.1)
Assessment Information
Assessors: Taylor, P1. Balona, J.2 & da Silva, J.M.3
Reviewer: Howard, A.1
Institutions: 1University of the Free State, 2Gauteng and Northern Regions Bat Interest Group, 3South African National Biodiversity Institute
Previous Assessors and Reviewers: Jacobs, D., Cohen, L., Richards, L.R., Monadjem, A., Schoeman, C., MacEwan, K., Sethusa, T. & Taylor, P.
Previous Contributor: Child, M.F.
Assessment Rationale
This species has a very wide distribution globally occurring in the Palaearctic and the Afrotropics. Within the assessment region it has an extent of occurrence (EOO) of 205,022 km2. It is a rarely recorded, difficult to sample species with small subpopulations. Moreover, the species is severely fragmented with an overall suspected regional population of fewer than 2,000 individuals. There is no evidence of decline, therefore the species is listed as Near Threatened under the D1 criterion.
Regional population effects: As it is a short-winged species that is unable to disperse large distance it is unlikely to be experiencing immigration from neighbouring countries.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Taylor P, Balona J & da Silva JM. 2025. A conservation assessment of Rhinolophus blasii. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
The Peak-saddle Horseshoe Bat has a large range in the Palaearctic and the Afrotropics, throughout which it is widely but patchily distributed. In southern Africa, it occurs form northeastern South Africa and Eswatini, through Zimbabwe, Botswana and Mozambique. Its range extends through southern Malawi to East Africa, the Democratic Republic of the Congo, Somalia, Ethiopia and into North Africa. Altitude range is from sea level to 1,200 m.
The subspecies R. b. empusa is mostly restricted to southern Africa, including Malawi and Zambia, occurring marginally beyond in the eastern The Democratic Republic of Congo (Csorba et al. 2003) and Tanzania (Kock & Howell 1988). Regionally, it occurs in Eswatini (Monadjem 2005), Zimbabwe (Cotterill 1996), and there are isolated records from central and northern Mozambique (Monadjem et al. 2020). Within the assessment region, specifically, it occurs in Limpopo, Mpumalanga, and KwaZulu-Natal provinces of South Africa. Within Kruger National Park, it occurs in the Pafuri region. Historical records also exist from Gauteng Province, but it has not been collected there since 1956. It is widely but sparsely distributed in the eastern parts of the region. The southern African subspecies is not well represented in museums, with just over 60 specimens examined in Monadjem et al. (2020).
The extent of occurrence (EOO) within the assessment region, based on known recorded colonies, is 205,022 km2.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 0 m asl (Monadjem et al. 2024)
Elevation Upper Limit (in metres above sea level): 2,215 m asl (Monadjem et al. 2024)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
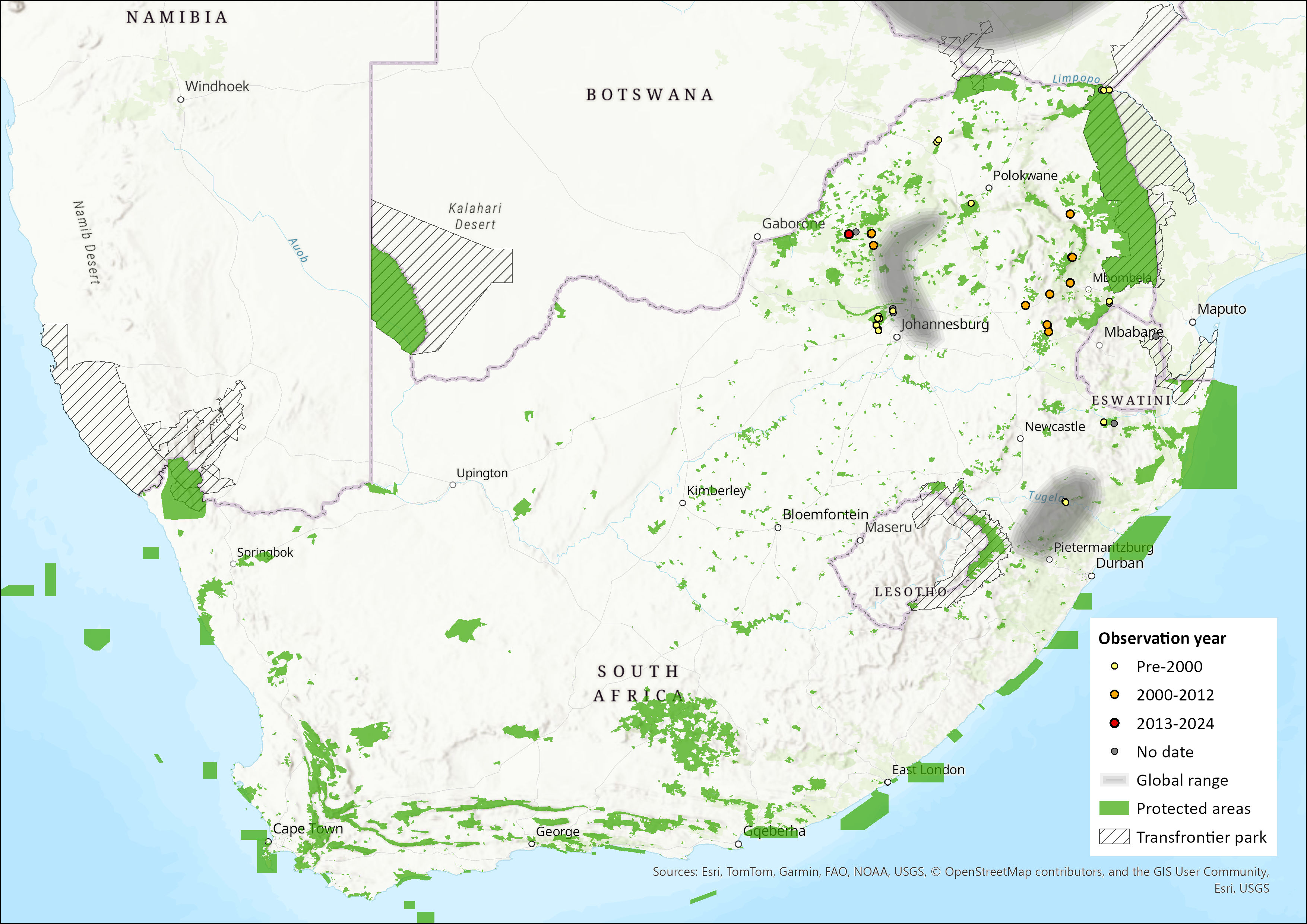 Figure 1. Distribution records for Peak-saddle Horseshoe Bat (Rhinolophus blasii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Peak-saddle Horseshoe Bat (Rhinolophus blasii) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical, Indomalayan, Palearctic
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Afghanistan | Extant | Native | – | – |
| Albania | Extant | Native | – | – |
| Algeria | Extant | Native | – | – |
| Armenia | Extant | Native | – | – |
| Austria | Extant | Native | – | – |
| Azerbaijan | Extant | Native | – | – |
| Bosnia and Herzegovina | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Bulgaria | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Croatia | Extant | Native | – | – |
| Cyprus | Extant | Native | – | – |
| Egypt | Presence Uncertain | Native | – | – |
| Eritrea | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Ethiopia | Extant | Native | – | – |
| Georgia | Presence Uncertain | Native | – | – |
| Greece | Extant | Native | – | – |
| Greece -> Kriti | Extant | Native | – | – |
| Iran, Islamic Republic of | Extant | Native | – | – |
| Israel | Extant | Native | – | – |
| Italy | Extinct Post-1500 | Native | – | – |
| Jordan | Extant | Native | – | – |
| Libya | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Montenegro | Extant | Native | – | – |
| Morocco | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| North Macedonia | Presence Uncertain | Native | – | – |
| Oman | Extant | Native | – | – |
| Pakistan | Extant | Native | – | – |
| Palestine, State of | Extant | Native | – | – |
| Romania | Presence Uncertain | Native | – | – |
| Russian Federation | Presence Uncertain | Native | – | – |
| Serbia | Extant | Native | – | – |
| Slovenia | Possibly Extinct | Native | – | – |
| Somalia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Syrian Arab Republic | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Tunisia | Presence Uncertain | Native | – | – |
| Turkmenistan | Extant | Native | – | – |
| Türkiye | Extant | Native | – | – |
| Yemen | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Most studies on the effects of climate change on bats are based on predictive species distribution modelling, thus there is a lack of empirical studies measuring behavioural, physiological, phenological or genetic responses to extreme and seasonal climatic changes, especially in the Global South (Festa et al. 2023; Pio et al. 2014). Globally, there have been documented declines in bat populations, species richness and distributions in relation to water availability with increasing global aridity which may become a growing concern as heat waves and maximum temperatures are expected to increase over much of South Africa (become a growing concern as heat waves and maximum temperatures are expected to increase over much of South Africa (Adams & Hayes 2021; Mbokodo et al. 2020).
Population information
In Africa, the species is not very common. Within the assessment region, it is rarely captured. However, this may be due to it having many small colonies. The overall population in the region is suspected to be low. It does not typically roost in large caves but prefers small caves, and mine adits where it is difficult to sample. There are only a small number of roost sites within the assessment region. Friedmann and Daly (2004) counted only four or five roosts. However, based on current distribution records, there are only between 20 and 30 major cave roosting sites within the assessment region, which we construe as separate subpopulations. There are thus suspected to be between 200 and 3,000 individuals in total.
Current population trend: Suspected to be stable
Continuing decline in mature individuals? Unknown
Number of subpopulations: 20-30
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: No
Number of mature individuals in largest subpopulation: 100
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
A phylogeographic study on the Rhinolophidae identified two clades within R. blasii in Africa, a northern clade including Iran, Algeria and Morrocco, and a southeastern clade compromised of individuals from Liberia, Tanzania, Malawi, Mozambique, Zimbabwe and South Africa. Within this southeastern clade, additional substructure is suggested involving South Africa, Mozambique and Zimbabwe (Demos et al. 2019), which could indicate a single metapopulation within the assessment region involving neighbouring countries; however, this would need to be validated with additional molecular work given that the study had limited sampling and only employed a single mitochondrial marker. Moreover, more finescale nuclear markers would be beneficial to specifically investigate genetic connectivity within the assessment region to confirm whether their distribution is indeed sparse and isolated, or whether simply a consequence of sampling/survey bias.
While population size estimates for the assessment region would indicate a low effective populations size (Ne) for the species (i.e., 20-900 assuming conversion ratios between 0.1-0.3), especially if multiple subpopulations exist, should the species actually form a metapopulation with neighbouring countries, its Ne might be adequate (e.g., >500). Focused population genetics research is needed to confirm this.
Habitats and ecology
In southern Africa, Peak-saddle Horseshoe Bats occur in savannah woodlands and are dependent on the availability of daylight roosting sites such as caves, mine adits or boulder piles (Skinner & Chimimba 2005). They are not always present in cave sites, which suggests that are partially migratory (Rautenbach 1982), but see Hutterer et al. (2005). Within the assessment region, the species is recorded from dolomitic geology and occurs in the Mopane Bioregion, Central Bushveld, Mesic Highveld Grassland.
They occur in groups of no more than three or four (Rautenbach 1982; Monadjem 2005; Skinner & Chimimba 2005). In Eswatini, they shared a roost (an abandoned gold mine) with several other bat species, including the numerically dominant R. clivosus (Monadjem 2005). It is a clutter forager and aerially hawks and gleans insect prey (Siemers & Ivanova 2004). Its diet consists mainly of Lepidoptera (Schoeman 2006).
This species is similar in size and appearance to other species in the region (e.g., R. simulator, and R. darlingi) but can be separated from these by the high-rising, sharply pointed connecting process characteristic of R. blasii and R. lobatus. The absence of axillary tufts of hair in the armpit distinguishes it from R. lobatus (Monadjem et al. 2020).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Suitable | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 3.8. Shrubland -> Shrubland – Mediterranean-type Shrubby Vegetation | – | Suitable | – |
| 7.1. Caves and Subterranean Habitats (non-aquatic) -> Caves and Subterranean Habitats (non-aquatic) – Caves | – | Suitable | – |
| 7.2. Caves and Subterranean Habitats (non-aquatic) -> Caves and Subterranean Habitats (non-aquatic) – Other Subterranean Habitats | – | Suitable | – |
| 8.2. Desert -> Desert – Temperate | – | Suitable | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Unknown
Congregatory: Unknown
Systems
System: Terrestrial
General Use and Trade Information
Not known to be traded.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
The major threat to this species within the assessment region is disturbance by tourist visits to caves, the use of the caves as shelters for livestock (ACR 2022), and the use of caves for traditional ceremonies by local communities. Re-commissioning of old mines is also a threat (Monadjem et al. 2020). The most recent record (J. Balona, unpubl. 2021) is from a cave threatened by blasting for a nearby development. Logging for firewood from local communities and deforestation is also suspected to be a threat.
It is unknown whether these threats are currently causing the population to decline within the assessment region.
Conservation
Protection of caves and monitoring of subpopulations is required. In the assessment region, the species is recorded from the protected areas: Kruger National Park; Wonderkop Nature Reserve; Cradle of Humankind Heritage Site (although no new records have been published from this site in decades), and Ithala Game Reserve. No specific interventions are necessary at present.
Recommendations for land managers and practitioners:
- Field surveys to discover new roost sites and confirm occupancy of existing roost sites.
Research priorities:
- Taxonomic research is needed to clarify the status of the African populations.
- Monitoring of population trends in response to the threat of mine closure or expansion, and renewable energy development, is recommended.
Encouraged citizen actions:
- Minimise disturbance at caves when visiting, and in particular, avoid hibernation and breeding periods.
- Citizens can report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP).
Bibliography
ACR. 2022. African Chiroptera Report 2022. AfricanBats, African Chiroptera Project, i-xix. Pretoria.
Adams, R.A. and Hayes, M.A. 2021. The importance of water availability to bats: climate warming and increasing global aridity. 50 years of bat research: foundations and new frontiers, pp.105-120.
Cotterill, F. P. D. 1996. New distribution records of insectivorous bats of the families Nycteridae, Rhinolophidae and Vespertilionidae (Microchiroptera: Mammalia) in Zimbabwe. Arnoldia Zimbabwe 10(8): 71-89.
Csorba, G.P., Ujhelyi, P. and Thomas, N. 2003. Horseshoe Bats of the World. Alana Books, Shropshire, England.
Demos, T.C., Webala, P.W., Goodman, S.M., Kerbis Peterhans, J.C., Bartonjo, M. and Patterson, B.D. 2019. Molecular phylogenetics of the African horseshoe bats (Chiroptera: Rhinolophidae): expanded geographic and taxonomic sampling of the Afrotropics. BMC Evolutionary Biology, 19(1), p.166.
Festa, F., Ancillotto, L., Santini, L., Pacifici, M., Rocha, R., Toshkova, N., Amorim, F., Benítez‐López, A., Domer, A., Hamidović, D. and Kramer‐Schadt, S. 2023. Bat responses to climate change: a systematic review. Biological Reviews, 98(1), pp.19-33.
Friedmann, Y. and Daly, B. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. CBSG Southern Africa, IUCN SSC Conservation Breeding Specialist Group, Endangered Wildlife Trust, South Africa.
Hutterer, R., Ivanova, T., Meyer-Cords, C. and Rodrigues, L. 2005. Bat migrations in Europe: a review of banding data and literature. 28, Naturschutz und Biologische Vielfalt. Federal Agency for Nature Conservation, Bonn, Germany.
Kock, D. and Howell, K.M. 1988. Three bats new for mainland Tanzania (Mammalia: Chiroptera). Senckenbergiana Biologica 68: 223–239.
Mbokodo, I., Bopape, M.J., Chikoore, H., Engelbrecht, F. and Nethengwe, N. 2020. Heatwaves in the future warmer climate of South Africa. Atmosphere, 11(7), p.712.
Monadjem A. 2005. Recording of the call of the Peak-saddle Horseshoe Bat (Rhinolophus blasii Peters, 1867) from Swaziland. African Bat Conservation News 3: 5–6.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2020. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis, 2nd Ed., University of Witwatersrand Press, Johannesburg.
Pio, D.V., Engler, R., Linder, H.P., Monadjem, A., Cotterill, F.P., Taylor, P.J., Schoeman, M.C., Price, B.W., Villet, M.H., Eick, G. and Salamin, N. 2014. Climate change effects on animal and plant phylogenetic diversity in southern Africa. Global Change Biology, 20(5), pp.1538-1549.
Rautenbach, I.L. 1982. Mammals of the Transvaal. No. 1, Ecoplan Monograph, Pretoria, South Africa.
Schoeman, M.C. 2006. The relative influence of competition and coevolution on the community structure of insectivorous bats in southern Africa. Ph.D. Thesis. University of Cape Town.
Sherwin, H.A., Montgomery, W.I. and Lundy, M.G. 2013. The impact and implications of climate change for bats. Mammal Review 43: 171–182.
Siemers, B.M. and Ivanova, T. 2004. Ground gleaning in horseshoe bats: comparative evidence from Rhinolophus blasii, R. euryale and R. mehelyi. Behavioral Ecology and Sociobiology 56: 464– 471.
Skinner, J.D. and Chimimba, C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.