

Malagasy Free-tailed Bat
Tadarida fulminans

2025 Red list status
Data Deficient
Regional Population Trend
Unknown
Change compared
to 2016
Changed
Overview
Tadarida fulminans – (Thomas, 1903)
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – MOLOSSIDAE – Tadarida – fulminans
Common Names: Malagasy Free-tailed Bat, Large Free-tailed Bat, Large Guano Bat, Lightning Guano Bat, Madagascan Large Guano Bat, Madagascan Large Free-tailed Bat, Madagascar Large Free-tailed Bat (English), Madagaskarse Groot Losstertvlermuis (Afrikaans)
Synonyms: Tadarida mastersoni Roberts, 1946
Taxonomic Note:
The African mainland population of the Malagasy Free-tailed Bat is geographically separated from the Madagascar population, and although these populations show overlap in cranial and external morphological characteristics, they may ultimately be identified as separate species (Goodman & Cardiff 2004; Monadjem et al. 2020). In this case, the mainland population could be reclassified Nyctinomus mastersoni (Roberts 1946) (Monadjem et al. 2020). Currently, no subspecies of T. fulminans are recognised (Skinner & Chimimba 2005). The three large Tadarida species are easily distinguished from other large molossid species by their unwrinkled upper lips (Monadjem et al. 2020).
Red List Status
Red List Status: DD – Data Deficient (IUCN version 3.1)
Assessment Information
Assessors: Richards, L.R.1, Taylor, P.J.2 & da Silva, J.M.3
Reviewer: Richardson, E.J.4
Institutions: 1Durban Natural Science Museum, 2University of the Free State, 3South African National Biodiversity Institute, 4Independent Consultant at Richardson & Peplow Environmental
Previous Assessors and Reviewers: Monadjem, A., Schoeman, C., Cohen, L., Jacobs, D., MacEwan, K., Richards, L, Sethusa, T. & Taylor, P.
Previous Contributors: Page-Nicholson, S., Relton, C. & Raimondo, D.
Assessment Rationale
Although the species has a very restricted range within the assessment region, being recorded only from the Pafuri region of Kruger National Park (KNP), it is widespread (although patchily distributed) elsewhere in Zimbabwe and East Africa. Because it occurs exclusively in a protected area, and there are no plausible threats within the assessment region or throughout its range, it does not qualify for Vulnerable D2. Its population size is unknown but is assumed to be stable inside KNP. We list this species as Data Deficient, owing to the paucity of data relating the biology, ecology and population size within the assessment region.
Regional population effects: It has a high wing loading (Monadjem et al. 2020), and thus we assume dispersal and rescue effects are possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: The species has not been recorded in recent times and there is limited information on the general biology, ecology, and population within the assessment region. Further research is needed to determine the presence, and conservation status, of the species within the assessment region.
Red List Index
Red List Index: Changed
Regional Distribution and occurrence
Geographic Range
This species ranges through East Africa, southern Africa and a few localities on the island of Madagascar (Monadjem et al. 2016). It has been recorded from a number of regions in eastern and southeastern Africa, along the border of Kenya and Uganda, as far south as Zimbabwe, central and northern Mozambique, southern Malawi, the very northern reaches of South Africa, with an isolated record in northeast Zambia (Monadjem et al. 2020, 2016). Specimens from eastern Democratic Republic of the Congo form the most westerly records (Monadjem et al. 2020). In Madagascar, this species is predominantly restricted to the central-southern region near to Fianarantsoa and Isalo National Park, and there are records from Tolagnaro near the southeast coast (Jenkins et al. 2007; Cotterill et al. 2008). Its altitudinal range extends from about sea level (at Fort Dauphin in Madagascar) to close to 2,000 m asl (in the Albertine Rift of East Africa). In the assessment region, the species is recorded only from the northern areas of the Limpopo Province of South Africa in Pafuri, KNP. The estimated extent of occurrence is 194 km².
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 0 m asl
Elevation Upper Limit (in metres above sea level): 2,000 m asl
Depth Lower Limit (in metres below sea level): N/A
Depth Upper Limit (in metres below sea level): N/A
Depth Zone: N/A
Map
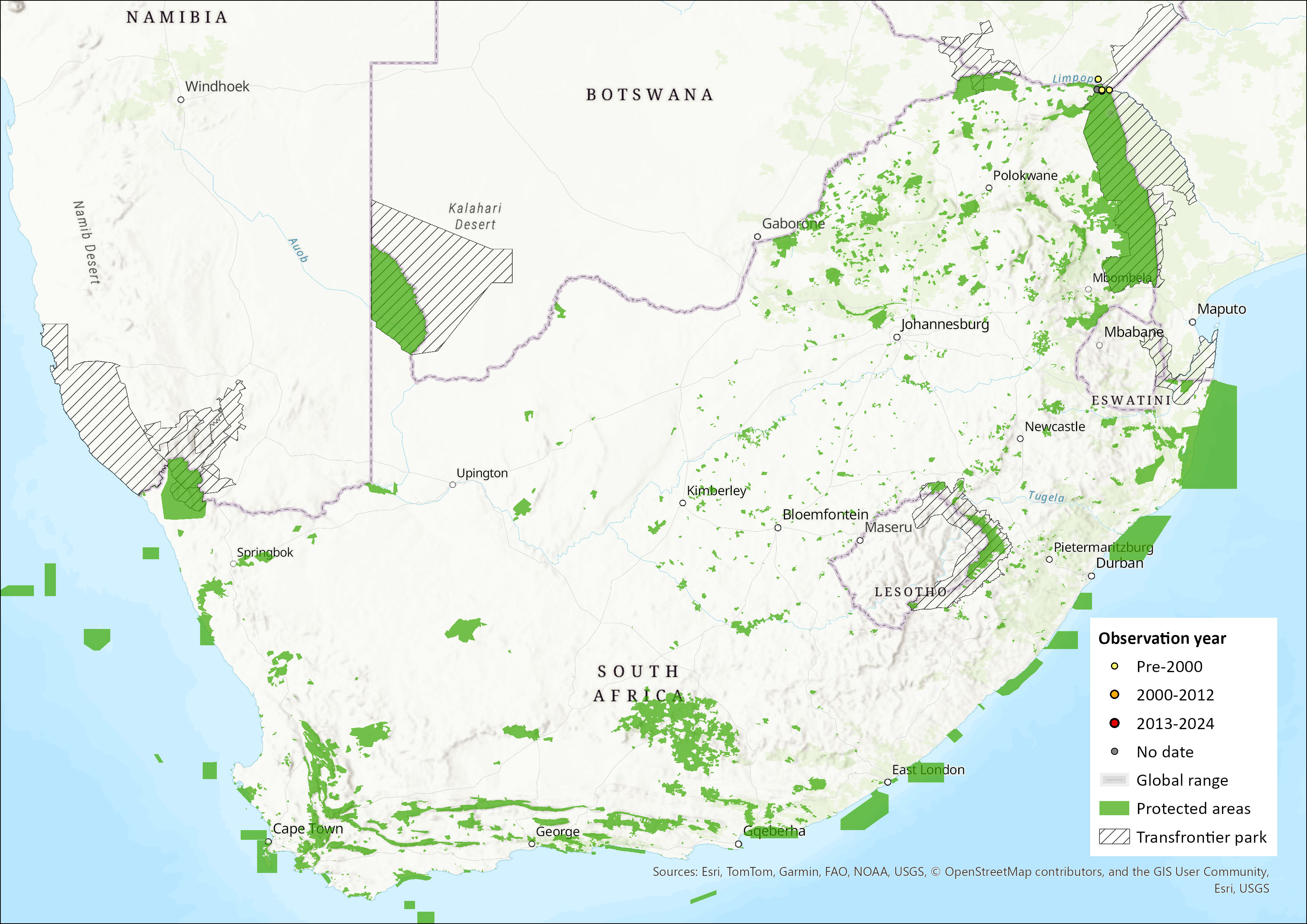 Figure 1. Distribution records for Malagasy Free-tailed Bat (Tadarida fulminans) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Malagasy Free-tailed Bat (Tadarida fulminans) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Madagascar | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | Resident |
| Rwanda | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: N/A
FAO Area Occurrence
FAO Marine Areas: N/A
Climate change
Limited occurrence records preclude species distribution and predictive niche modelling. Understanding the limiting bioclimatic or ecogeographic variables and the potential impacts of climate change on the species is therefore difficult. Notwithstanding, the species is associated with both moist and dry savanna characterised by rocky outcrops and inselbergs (Cotterill and Ferguson 1993a). It is therefore likely that increased drought frequency in dry savannas may impact the vegetation and insect prey base (likely to be moths as is the case for similar-sized large molossids) (Gaona et al. 2024). This is turn may result in an altered distribution pattern within the assessment region.
Population information
Although sporadically distributed in mainland Africa, it is considered locally common in the rest of its range. In southern Africa, it roosts communally in small to medium-sized groups, which may number over 30 individuals (Cotterill 2001), but do not exceed 100 individuals (ACR 2024; Monadjem et al. 2016).
Current population trend: Unknown
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individuals in one subpopulation: Assumed to comprise one population based on wing loading and presumed dispersal capabilities
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Likely only one subpopulation in the assessment region
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: Unknown
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: Unknown
Probability of extinction in the wild within 100 years: Unknown
Population genetics
To date, only a broadscale phylogenetic study looking at the evolutionary history of free-tailed bats in the subfamily Molossinae has been conducted involving this species (Ammermann et al. 2012). Within the assessment region, it is localised to the very north of South Africa, and it is expected to form a single genetically distinct subpopulation within the global extent of the species; however, this needs confirmation through a more comprehensive population genomic study.
Habitats and ecology
The Malagasy Free-tailed Bat is typically a savannah species; found in both dry and moist savannah habitats (Monadjem et al. 2016). It is not thought to be dependent on caves, but is found in rocky areas, such as basalt, dolomite and especially granite outcrops with crevices, which are common throughout southern and eastern Zimbabwe (Cotterill 1996). This species is commonly associated with miombo and mopane savannah woodlands, interspersed with steep-sided mountains ridges or inselbergs (Cotterill 2001; Monadjem et al. 2020). In the assessment region, the species is recorded from the Lowveld and Mopane bioregions.
It is a communal rooster and roosts in crevices in vertical rock faces, which may occur more than 60 m above the ground (Cotterill & Fergusson 1993a) and in spaces under exfoliating rock (Cotterill 2001). It is an open-air forager. Together with T. ventralis, the Malagasy Free-tailed Bat has the longest, narrowest wing tips of all molossid bat species (Monadjem et al. 2020), which is an adaptation for fast and agile flight. In southern Africa, there is no information on the diet of this species (Skinner & Chimimba 2005; Monadjem et al. 2020). However, in Sengwa (Zimbabwe) the species diet consists of Lepidoptera and Coleoptera (Fenton, 1985).
Ecosystem and cultural services: None recorded.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Marginal | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 6. Rocky areas (eg. inland cliffs, mountain peaks) | – | Suitable | – |
Life History
Generation Length: Assumed to be 2-3 years
Age at Maturity: Female or unspecified: Likely 1-2 years
Age at Maturity: Male: Likely 1-2 years
Size at Maturity (in cms): Female: Mean total length = 15.60 ±0.37 cm; Mean forearm length = 6.02 ±0.13 cm (Monadjem et al. 2020)
Size at Maturity (in cms): Male: Mean total length = 15.10 ±1.41 cm; Mean forearm length = 5.98 (Monadjem et al. 2020)
Longevity: Unknown
Average Reproductive Age: Likely 1-2 years
Maximum Size (in cms): Unknown
Size at Birth (in cms): Unknown
Gestation Time: (Not specified) 1100 – 110 days (Cotterill and Ferguson 1993b)
Reproductive Periodicity: Females from Zimbabwe were seasonally polyoestrous, giving birth during the austral summer and early winter (Cotterill and Ferguson 1993b).
Average Annual Fecundity or Litter Size: one pup
Natural Mortality: Not specified
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Unknown
Congregatory: Assumed to be congregatory
Systems
System: Terrestrial
General Use and Trade Information
There is no evidence to suggest that this species is traded or harvested within the assessment region.
Local Livelihood: N/A
National Commercial Value: N/A
International Commercial Value: N/A
End Use: N/A
Is there harvest from captive/cultivated sources of this species? N/A
Harvest Trend Comments: N/A
Threats
Across its range there are no major threats to this species (ACR 2024; Monadjem et al. 2016). Considering it occurs exclusively within a protected area (KNP) in the assessment region, no plausible threats have been identified.
Conservation
There are currently no active conservation measures necessary for this species within the subregion as it occurs exclusively within the protected Kruger National Park in South Africa.
Research priorities:
- Additional studies into the geographic range and taxonomic status of individuals allocated to T. fulminans are required (Monadjem et al. 2016), and particularly the relationship between African mainland and Malagasy populations needs to be investigated.
- Targeted surveys within likely roosting areas (e.g. rocky outcrops and inslebergs with crevices) are needed to determine its occurrence beyond that of the known distribution.
Encouraged citizen actions:
- Citizens can assist the conservation of the species by reporting sightings on virtual museum platforms (for example, iNaturalist and MammalMAP) and therefore contribute to an understanding of the species distribution.
- Any downed bats must be reported to the nearest Bat Interest Group or museum.
Bibliography
ACR. 2024. African Chiroptera Report 2023. Van Cakenberghe, V. and Seamark, E.C.J. (Eds.) African Chiroptera Project, Pretoria. i – xviii; 1 – 1210 pp.
Ammerman, L. K., Lee, D.N. and Tipps, T.M. 2012. First molecular phylogenetic insights into the evolution of free-tailed bats in the subfamily Molossinae (Molossidae, Chiroptera). Journal of Mammalogy, 93 (1): 12 – 28. doi: http://dx.doi.org/10.1644/11-MAMM-A-103.1
Cotterill, F.P.D. and Fergusson, R.A. 1993q. Capturing free-tailed bats (Chiroptera: Molossidae): the description of a new trapping device. Journal of Zoology 231: 645–651.
Cotterill, F.P. and Fergusson, R.A. 1993b. Seasonally polyestrous reproduction in a free-tailed bat Tadarida fulminans (Microchiroptera: Molossidae) in Zimbabwe. Biotropica 1:487-92.
Cotterill, F.P.D., Hutson, A.M., Racey, P.A. and Ravino, J. 2008. Tadarida fulminans. Page e.T21316A9269805. The IUCN Red List of Threatened Species.
Cotterill, F.P.D. 1996. New distribution records of free-tailed bats (Microchiroptera: Molossidae) in Zimbabwe. Arnoldia Zimbabwe 10: 91–102.
Cotterill, F.P.D. 2001. Further notes on large Afrotropical free-tailed bats of the genus Tadarida (Molossidae: Mammalia). Arnoldia Zimbabwe 10: 199–210.
Fenton, M.B. 1985. The feeding behaviour of insectivorous bats: echolocation, foraging strategies, and resource partitioning. Transvaal Museum Bulletin, 21(5).
Gaona, F.P., Delabye, S., Potocky, P., Govorov, V., Cuda, J., Foxcroft, L.C., Garlacz, R., Hejda, M., MacFadyen, S., Pyrcz, T, and Pyskova, K. 2024. Climate-driven vegetation characteristics shape phytophagous and carnivorous insect diversity in South African savannahs. bioRxiv. 2024:2024-07.
Goodman, S.M. and Cardiff, S.G. 2004. A new species of Chaerephon (Molossidae) from Madagascar with notes on other members of the family. Acta Chiropterologica 6: 227–248.
Jenkins, R.K.B., Kofoky, A.F, Russ, J.M., Friafidison, A., Siemers, B.M., Randrianandrianina, F.H., Mbohoahy, T., Rahaingodrahety, V.N and Racey, P.A. 2007. Ecology of Bats in the southern Anosy Region. In: J. U. Ganzhorn, S. M. Goodman, and M. Vincelette (eds), Biodiversity, ecology and conservation of littoral ecosystems in southeastern Madagascar, Tolagnaro (Fort Dauphin), Smithsonian Institution, Washington, DC, USA.
Monadjem, A., Ravino, J., Hutson, A.M., Cotterill, W. and Racey, P.A. 2016. Tadarida fulminans. The IUCN Red List of Threatened Species 2017: e.T21316A22122012.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2020. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis, second edition.University of Witwatersrand Press, Johannesburg.
Skinner, J.D. and Chimimba, C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.