

Lesser Red Musk Shrew
Crocidura hirta

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Crocidura hirta – Peters, 1852
ANIMALIA – CHORDATA – MAMMALIA – EULIPOTYPHLA – SORICIDAE – Crocidura – hirta
Common Names: Lesser Red Musk Shrew (English), Klein Rooiskeerbek (Afrikaans)
Synonyms: No Synonyms
Taxonomic Note:
Four subspecies are listed (Heim de Balsac & Meester 1977), of which two occur in southern Africa: C. c. hirta, found in the wetter eastern areas of the country with annual rainfall over 500 mm, occurring in KwaZulu-Natal, Free State, North West, Mpumalanga and Limpopo provinces, as well as Zimbabwe and Mozambique; while C. h. deserti (the arid variant) occurs in the drier western regions in northern Northern Cape, northern Botswana and north-eastern Namibia (Meester et al. 1986) Further research should resolve the taxonomy of these variants as there may be more than one species across the range.
Red List Status: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Russo, I.M.1, da Silva, J. M.2
Reviewer: Erusan, R.3
Institutions: 1Cardiff University, 2South African National Biodiversity Institute, 3South African National Parks
Previous Assessors: Taylor, P., Baxter, R. & Monadjem, A.
Previous Reviewer: Child, M.F.
Previous Contributors: Avery, M., MacFadyen, D., Avenant, N., Wilson, B. & Palmer, G.
Assessment Rationale
This is a widely distributed and common species within the assessment region, occurring in numerous protected areas (including the Kruger National Park, Golden Gate Highlands National Park, and presumably the Kgalagadi Transfrontier Park). It has a broad habitat tolerance across its range, including agricultural landscapes (as long as they are not overgrazed) and gardens, and is one of the most common Crocidura species in suitable habitat. As it is largely commensal with humans, landscape transformation and modification areunlikely to have a negative effect. The species is therefore listed as Least Concern. Similarly to other shrew species interventions such as protected area expansion of moist grassland habitats, as well as incentivising landowners to sustain natural vegetation around wetlands and livestock or wildlife at ecological carrying capacity to avoid overgrazing should be incorporated into management.
Regional population effects: This species is too small to disperse over long distances so no significant rescue is possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Russo IM da Silva JM. 2025. A conservation assessment of Crocidura hirta. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This is one of the most widespread shrew species in Africa, occurring throughout Central, East and southern Africa (Skinner and Chimimba 2005). Within the assessment region, they occur in all provinces except the Western and Eastern Cape, and only sparsely in the Mpumalanga province, with the southern limit around Pennington in the KwaZulu-Natal Province. Similarly, it occurs throughout the Middleveld, Lowveld and Lubombo regions, and marginally in the Highveld region at Mbabane, in Eswatini (Monadjem 1998). Although Barn Owl (Tyto alba) pellet analyses indicate a wide presence in the Northern Cape Province (records from Spitzkop Nature Reserve (Avery and Avery 2011), there is morphological and distributional overlap with C. cyanea and thus the mandibles retrieved from Barn Owl pellets could be confused between the two species (Taylor and Contrafatto 1996). Meester (1963) has confirmed the presence of the species in the Kalahari but more vetting of museum specimens and Barn Owl pellets is needed to confirm the Karoo and West Coast distribution. A recent record from Goegap Nature Reserve record would mean a considerable range extension to the west (Avery and Avery 2011). It has been confirmed to occur widely in the bushveld and Kalahari regions of the North West Province during a recent survey (Power 2014).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
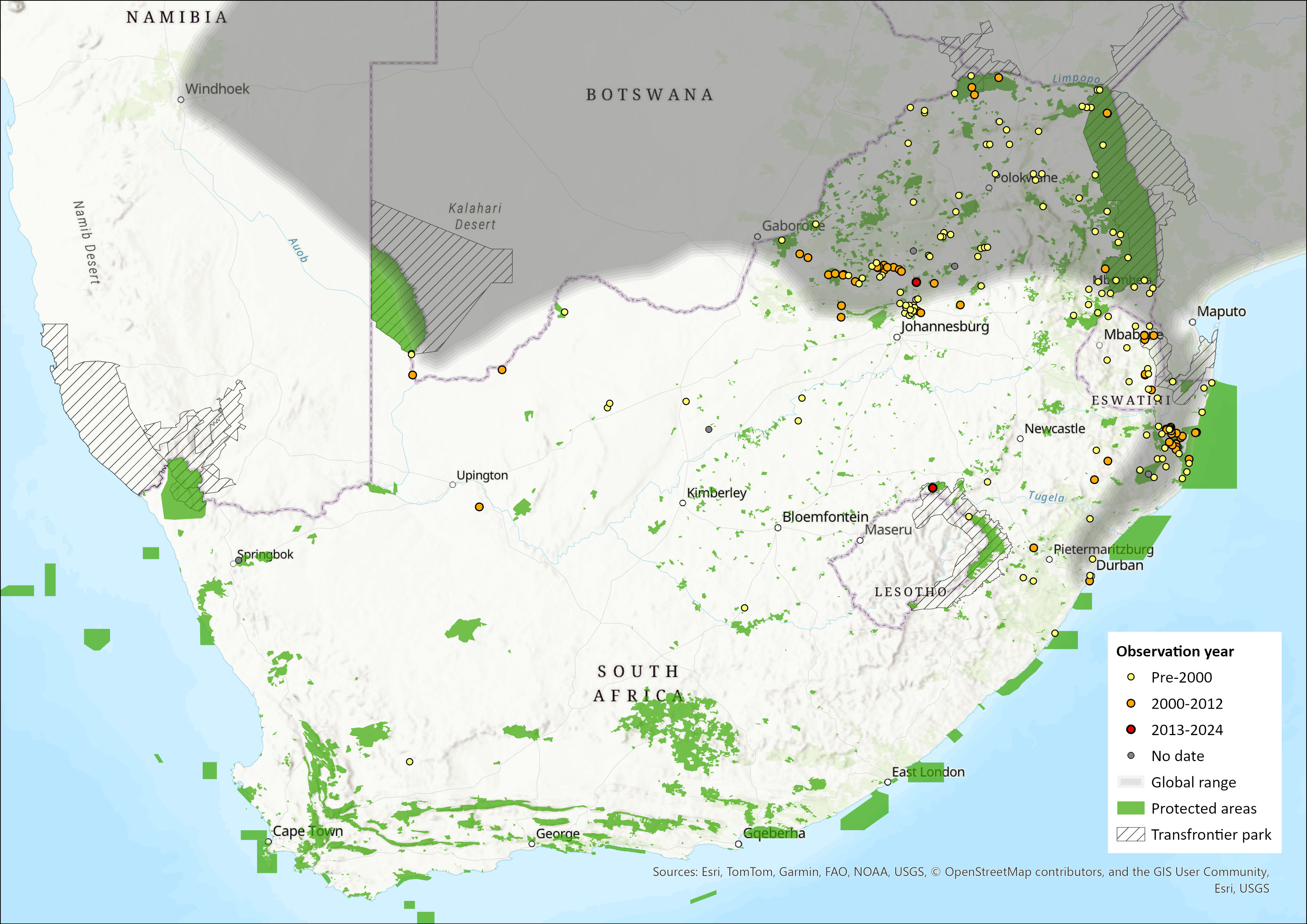
Figure 1. Distribution records for Lesser Red Musk Shrew (Crocidura hirta) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Kenya | Presence Uncertain | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Presence Uncertain | Native | – | – |
| Somalia | Presence Uncertain | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Climate change is not predicted to become a major threat for this species as savannahs are projected to expand (for example, Kgope et al. 2010).
Population
This is one of the most common shrew species in the assessment region and it the only shrew commonly recorded from the Lowveld region, where it exhibits seasonal fluctuations in population size following fire (Kern 1981), population increases are generally related to successional regrowth of vegetation and grass cover where a crash is observed after fire has removed the undergrowth. It is frequently recorded in the northern bushveld regions of the North West Province (Power 2014). In Mkhuze Game Reserve, KwaZulu-Natal Province, C. hirta and C. fuscomurina accounted for 73% of all shrew captures, where C. hirta was the most abundant (Delcros et al. 2014). Similarly, at Phinda Private Game Reserve, KwaZulu-Natal Province, C. hirta and C. fuscomurina were the most frequently captured shrews across all habitats but C. fuscomurina was slightly more abundant. In Golden Gate Highlands National Park, C. hirta was the least common species with C. cyanea and C. flavescens being more dominate. This species can be dominant in certain habitats. Furthermore, it is well adapted to living in association with humans or in habitats modified by humans and can be common in gardens.
In an ungrazed natural grassland near Matshapa (Eswatini) density ranged between 0.7±.4 and 2.5±.9 individuals/ha between 1995 and 1998 (Monadjem and Perrin 2003).
Population Information
Current population trend: Stable. Occurs in a wide variety of habitats and in both urban and agricultural habitats, so no reason to suspect population decline based on habitat loss.
Continuing decline in mature individuals: No
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: No. Occurs in a wide variety of habitats and in both urban and agricultural habitats.
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: (Not specified)
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
This species has been investigated in a phylogenetic context using mitochondrial and ddRAD data (Dianat et al. 2023). While not a comprehensive look at the population genetic structure within this species, Dianat et al. (2023) did find substantial cryptic diversity within C. hirta, namely within Tanzania, Mozambique and Zambia. It is possible further subdivisions may be found within the assessment region pending a more comprehensive examination. Given the limited information on population structure and the lack of population size estimates, the effective population size (Ne) for this species cannot be quantified.
Habitats and ecology
This species is found in habitats such as grassland, savannah, scrublands and compost heaps in gardens. In the eastern parts of its range, it is found primarily in moist habitats along streams and wetlands and requires cover through dense undergrowth, shrubs, disused rodent burrows, termitaria holes or beneath rocks (Skinner and Chimimba 2005). In the North West Province, it was taken from a variety of habitats such as vleis and dense bushveld with rank grass (Power 2014). In the KwaZulu-Natal Province, they have been collected from grass clumps, reedbeds, fig-dominated forests, savannah woodland, tall grassland and fallow agricultural fields (Taylor 1998). In Mhkuze and Phinda Game Reserves, it was found in woodland, floodplain grassland, Lebombo thicket and sand forest habitat (Delcros et al. 2014, Rautenbach et al.2014). In Swaziland the species occurs in a wide range of vegetation types from moist Middleveld grasslands to Acacia and broad-leaf savanna in the Lowveld region but the species is closely associated with areas which have tall, dense grass and absent from Highveld sour grassland (Monadjem 1998). In its western range, individuals occur in scrub bushes and riverine woodland where there is debris. They probably rely on dew in the water-scarce Kalahari. They show a preference for dense vegetation, deep leaf litter and proximity to water (Dickman 1995, Rautenbach et al. 2014). In the Maluti Mountains they prefer steep slopes with high soil moisture, to escape predation from black blacked jackals and domestic cats.
It is commensal with humans and occurs in modified landscapes, such as gardens and agricultural areas. It thus fulfils a similar niche to C. flavescens except occurring more in the north of the assessment region. In Eswatini, for example, C. hirta is found in the Lowveld whereas C. flavescens is found at higher altitudes (Monadjem 1999), which corresponds broadly with a savannah versus grassland niche respectively. Both these species have been found in the Maluti Mountains (R. Erusan, pers. comm. 2025). This area has extreme temperatures including snow and heat waves.
They are active throughout the day and night but predominantly nocturnal (Baxter et al. 1979). The species is generally solitarily but Meester (1963) showed they can live together in pairs and groups. While mainly insectivorous, small amounts of plant material are also ingested (Dickman 1995). They also feed on carcasses of rodents and bats and can be cannibalistic (Meester 1963).
Ecosystem and cultural services: An important prey species (e.g., Avery and Avery 2011).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 4.5. Grassland -> Grassland – Subtropical/Tropical Dry | – | Suitable | – |
| 8.1. Desert -> Desert – Hot | – | Suitable | – |
| 14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land | – | Unknown | – |
| 14.2. Artificial/Terrestrial -> Artificial/Terrestrial – Pastureland | – | Unknown | – |
| 14.3. Artificial/Terrestrial -> Artificial/Terrestrial – Plantations | – | Unknown | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Unknown | – |
| 14.6. Artificial/Terrestrial -> Artificial/Terrestrial – Subtropical/Tropical Heavily Degraded Former Forest | – | Unknown | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
There is no known subsistence or commercial use of this species.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
The main threat to shrews is the loss or degradation of moist, productive areas such as wetlands and rank grasslands within suitable habitat. The two main drivers behind this are abstraction of surface water and draining of wetlands through industrial and residential expansion, and overgrazing of moist grasslands, which leads to the loss of ground cover and decreases small mammal diversity and abundance (Bowland and Perrin 1989, 1993; Monadjem 1999). Suppression of natural ecosystem processes, such as severe fire can also lead to habitat degradation through bush encroachment or loss of plant diversity through alien invasives and is suspected to be increasing with human settlement expansion. There are also clear overlaps and synergistic effects between these threats. Based on loss of natural habitat, we infer a continuing decline in some local populations which may consequently lead to a progressive impact on the national population, should habitat loss continue. Natural predators also actively influence the population.
Current habitat trend: As this is predominantly a savannah species, it does not suffer as much from habitat loss as grassland or forest specialist species, as savannah remains relatively intact within the assessment region (Driver et al. 2012). Furthermore, as it commensal with humans, urban and rural expansion has fewer negative effects. Similarly, climate change is not predicted to become a major threat for this species as savannahs are projected to expand (for example, Kgope et al. 2010). Thus, the habitat for this species is stable.
Conservation
This species is found in several protected areas across its range which includes the Kruger National Park, Golden Gate Highlands National Park and the Kgalagadi Transfrontier Park. Although no interventions are currently necessary, protecting and restoring suitable habitat, such as moist grassland patches will benefit this species. Biodiversity stewardship schemes should be promoted to conserve such patches. Importantly, at the local scale, landowners and managers should be educated, encouraged and incentivised to conserve the habitats on which shrews and small mammals depend. Retaining ground cover is the most important management tool to increase small mammal diversity and abundance. This can be achieved through lowering grazing pressure (Bowland and Perrin 1989), or by maintaining a buffer strip of natural vegetation around wetlands (Driver et al. 2012). Small mammal diversity and abundance is also higher in more complex or heterogeneous landscapes, where periodic burning is an important tool to achieve this (Bowland and Perrin 1993). Removing alien vegetation from watersheds, watercourses and wetlands is also an important intervention to improve flow and water quality, and thus habitat quality, for shrews. Education and awareness campaigns should be employed to teach landowners and local communities about the importance of conserving wetlands and moist grasslands.
Recommendations for land managers and practitioners:
- Landowners and communities should be incentivised to stock livestock or wildlife at ecological carrying capacity to avoid overgrazing and to maintain a buffer of natural vegetation around wetlands.
- Enforce regulations on developments that can potentially impact on the habitat integrity of grasslands and wetlands.
Research priorities:
- Additional field surveys are needed to clarify and confirm the distribution of this species.
- Museum records must be confirmed to refine the distribution map.
- Molecular research is needed to revise the taxonomic status of putative subspecies.
Encouraged citizen actions:
- Citizens are requested to submit any shrews killed by cats or drowned in pools to a museum or a provincial conservation authority for identification, thereby enhancing our knowledge of shrew distribution (carcasses can be placed in a ziplock bag and frozen with the locality recorded).
- Practice indigenous gardening to sustain small mammals.
Bibliography
Avery, D.M. and Avery, G. 2011. Micromammals in the Northern Cape Province of South Africa, past and present. African Natural History 7: 9-39.
Baxter RM, Goulden EA, Meester J. 1979. The activity patterns of some southern African Crocidura in captivity. Acta Theriologica 24: 61-68.
Bowland, A.E. and Perrin, M.R. 1989. The effect of overgrazing on the small mammals in Umfolozi Game Reserve. Zeitschrift für Säugetierkunde 54: 251–260.
Bowland, J.M. and Perrin, M.E. 1993. Wetlands as reservoirs of small-mammal populations in the Natal Drakensberg. South African Journal of Wildlife Research 23: 39–43.
Delcros, G., Taylor, P.J., and Schoeman, M.C. 2015. Ecological correlates of small mammal assemblage structure at different spatial scales in the savannah biome of South Africa. Mammalia 79(1): 1-14.
Dickman CR. 1995. Diets and habitat preferences of three species of crocidurine shrews in arid southern Africa. Journal of Zoology 237: 499-514.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., Van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An assessment of South Africa’s biodiversity and ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Heim de Balsac, H. and Meester, J. 1977. Order Insectivora. In: J. Meester and H.W. Setzer (eds), The Mammals of Africa: An Identification Manual, pp. 1-29. Smithsonian Institution Press, Washington, D. C., USA.
Kern NG. 1981. The influence of fire on populations of small mammals of the Kruger National Park. Koedoe 24: 125-157.
Kgope BS, Bond WJ, Midgley GF. 2010. Growth responses of African savanna trees implicate atmospheric [CO2] as a driver of past and current changes in savanna tree cover. Austral Ecology 35: 451-463.
Meester, J. 1963. A systematic revision of the shrew genus Crocidura in Southern Africa. Transvaal Museum Memoir 13: 1-127.
Monadjem A, Perrin M. 2003. Population fluctuations and community structure of small mammals in a Swaziland grassland over a three-year period. African Zoology 38: 127-137.
Monadjem A. 1998. The mammals of Swaziland. Conservation Trust of Swaziland and Big Games Parks, Mbabane, Swaziland.
Monadjem A. 1999. Geographic distribution patterns of small mammals in Swaziland in relation to abiotic factors and human land-use activity. Biodiversity & Conservation 8: 223-237.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Rautenbach A, Dickerson T, Schoeman MC. 2014. Diversity of rodent and shrew assemblages in different vegetation types of the savannah biome in South Africa: no evidence for nested subsets or competition. African Journal of Ecology 52: 30-40.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Taylor PJ, Contrafatto G. 1996. Mandible shape and size in three species of small musk shrews (Crocidura Wagler, 1832) from southern Africa. Mammalia 60: 753-766.
Taylor, P. 1998. The Smaller Mammals of KwaZulu-Natal. University of Natal Press, Pietermaritzburg, South Africa.