

Large-eared free-tailed bat
Otomops martiensseni

2025 Red list status
Near threatened
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Otomops martiensseni – (Matschie, 1897)
ANIMALIA – CHORDATA – MAMMALIA – CHIROPTERA – MOLOSSIDAE – Otomops – martiensseni
Common Names: Large-eared Free-tailed Bat, Large-eared Giant Mastiff Bat, Giant Mastiff Bat, Martiensen’s Free-tailed Bat, Martienssen Bat, Martienssen’s Big-eared Bulldog Bat (English), Bakoor-losstertvlermuis
(Afrikaans)
Synonyms: Otomops martiensseni (Chubb 1917) spp. Icarus, Nyctinomus martiensseni (Matschie 1897)
Taxonomic Note:
Two subspecies, including the nominate Otomops martiensseni martiensseni and O. m. icarus (from South Africa, Zimbabwe and Angola) were recognised by Meester et al. (1986). However, subsequent ecological, morphometric and molecular data demonstrated a single species within southern, western and eastern Africa, with ongoing gene flow between widely scattered populations (Lamb et al. 2006, 2008; Richards et al. 2012; Ralph et al. 2015). In light of a newly described northeastern species, O. harrisoni, the distribution of O. martiensseni in sub-Saharan Africa has been revised to exclude the Arabian Peninsula, Djibouti, Eritrea, and Ethiopia, (Ralph et al. 2015; Patterson et al. 2018). Although previously included within O. martiensseni (Simmons 2005), populations from Madagascar are now considered a distinct species (O. madagascariensis) following genetic and morphometric studies (Dorst 1953; Lamb et al. 2008; Richards et al. 2012; Ralph et al. 2015).
Red List Status: NT – Near Threatened, D1 (IUCN version 3.1)
Assessment Information
Assessors: Richards L.R.1, Naidoo, T.1 & da Silva J.M.2
Reviewer: Smith, C.3
Institutions: 1Durban Natural Science Museum, 2South African National Biodiversity Institute, 3Endangered Wildlife Trust
Previous Assessors and Reviewers: Richards, L.R., Schoeman, C., Taylor, P., White, W., Cohen, L., Jacobs, D., MacEwan, K., Sethusa, T. & Monadjem, A.
Previous Contributors: Child, M.F., Relton, C. & Raimondo, D.
Assessment Rationale
This species is widely but patchily distributed across much of sub-Saharan Africa. Within the assessment region, it has been recorded from several localities in KwaZulu-Natal around the eThekwini area and has recently been shown to be more widely distributed, having been recorded from Kruger National Park, Mapungubwe National Parks (Limpopo Province), and Mbotyi and Manubi State Forests (Eastern Cape Province). The estimated extent of occurrence is 258,823 km2. Subpopulations are suspected to be stable and (in Limpopo at least) fairly well protected in the assessment region. However, extant synanthropic roof roosts within the eThekwini region, on which the species relies, are rare and may still be subject to persecution from ill-informed homeowners. Of concern, very few breeding males have been recorded or re-sampled in eThekwini within the recent past (K. Richardson unpubl. data). While it is difficult to sample due to its high-altitude foraging behaviour, it is considered rare (typically < 1% of recorded calls during acoustic sampling; Adams et al. 2015, Moir et al. 2020). Since there are typically 10–30 mature individuals in each colony within the assessment region, we infer a mature population size of 330–990 animals. While continuing decline is not suspected, colony sizes are small and vulnerable to local extinction due to roost disturbance and loss. This species thus qualifies as Vulnerable D1 as inferred population size is < 1,000 mature individuals. However, due to its good dispersal capacity, we assume rescue effects are possible and employ the regional criterion to downlist to Near Threatened D1. Further data on its occurrence, subpopulation sizes and trends are urgently needed, and this species may qualify for a more threatened listing once additional data are available. Furthermore, O. martiensseni is considered a high-risk species for impacts / fatalities from collision with wind turbines or barotrauma (MacEwan et al. 2020). Although no fatalities have been reported yet, the expansion of the wind energy footprint within the Eastern Cape necessitates close monitoring for potential impacts to the species within the region.
Regional population effects: With the discovery of subpopulations in Limpopo and Botswana, this species’ range is suspected to be connected with that of Zimbabwe and northeastern Mozambique. Wing-loading is high (Norberg & Rayner 1987), and thus dispersal capacity is assumed to be good, indicating that rescue effects are possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No Change
Red List Index
Red List Index: No Change
Recommended citation: Richards LR, Naidoo T & da Silva JM. 2025. A conservation assessment of Otomops martiensseni. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The Large-eared Giant Mastiff Bat has a patchy distribution across Africa occurring from Ghana and Côte d’Ivore in the West, extending eastward through to Central African Republic, Rwanda, Uganda, Tanzania, Kenya and southwards towards Angola, Zambia, Zimbabwe, northwestern Mozambique and South Africa (Richards et al. 2012, Adams et al. 2015, Ralph et al. 2015, Patterson et al. 2018, Brinkley et al. 2019, Parker and Bernard 2018, Moir et al. 2020, Monadjem et al. 2020). Adams et al. (2015) also recorded its presence in the Tuli Block of Botswana (near the South African border), the first published records for the country, which indicates the species is more widespread in southern Africa than previously thought but the species continues to be rare throughout most of its range. Within the assessment region, this species was originally thought to be restricted to several localities in the KwaZulu-Natal Province, centred around eThekwini Municipality (Fenton et al. 2004; Monadjem et al. 2020). However, acoustic data has expanded its range 870 km north to Limpopo Province where the species has recently been recorded from several localities in Kruger National Park (KNP) and Mapungubwe National Parks (MNP) (Adams et al. 2015; Parker & Bernard 2018). It has also recently been acoustically (G. De Jong, unpublished data) recorded from Mpumalanga. These data thus bridge the gap between the most southerly localities in Mbotyi and Manubi State Forests along the coastal Eastern Cape region (Moir et al. 2020), with those of the nearest known occurrence records from southwestern Zimbabwe in 1978 at the Sengwa Wildlife Research Area (Fenton & Bell 1981), and northwestern Zimbabwe near the Kariba Dam (Hutton 1986). Current gaps in distribution probably reflect insufficient sampling rather than absence. Current estimated extent of occurrence within the assessment region is 258,823 km².
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 0
Elevation Upper Limit (in metres above sea level): 2900
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
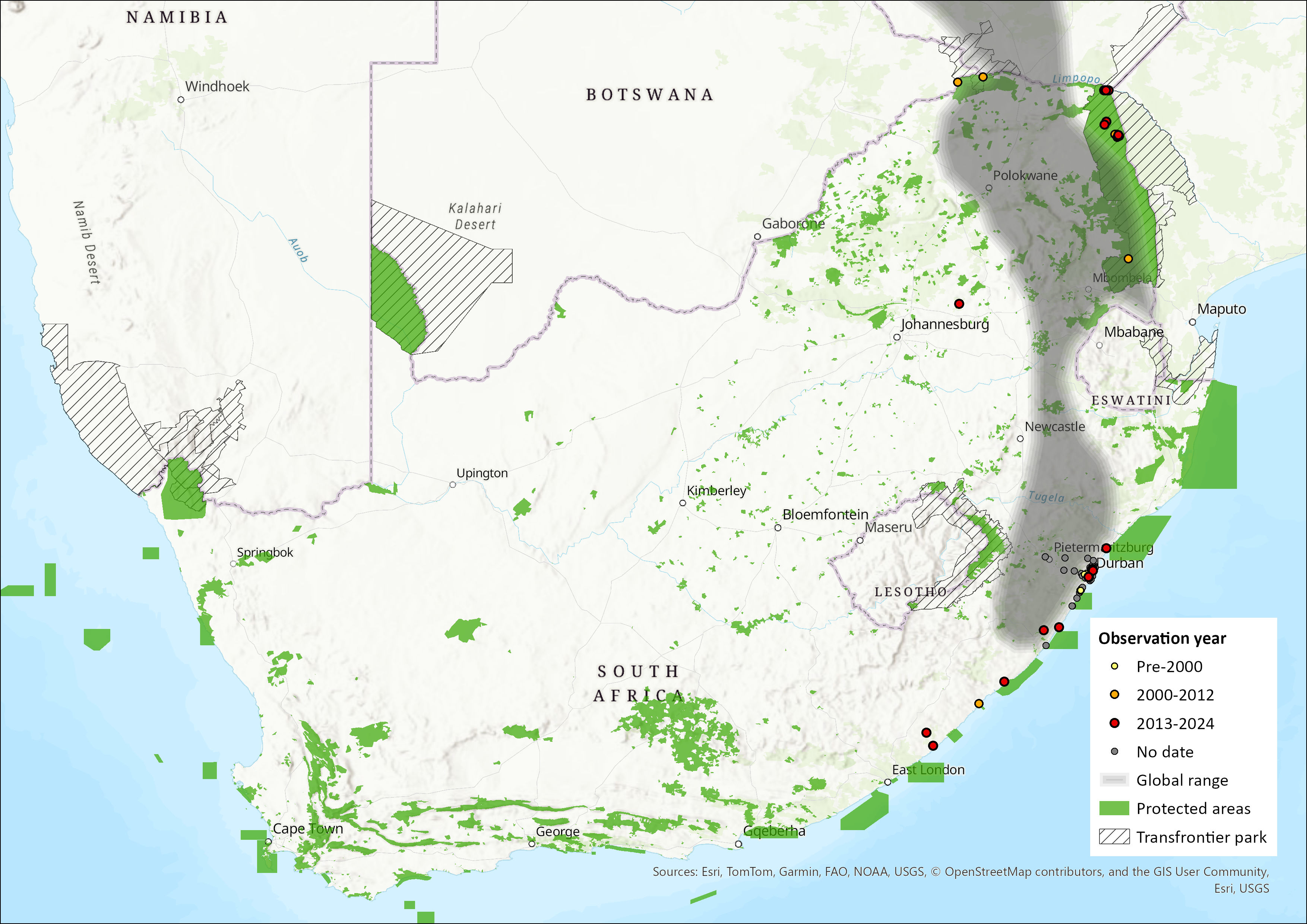 Figure 1. Distribution records for Large-eared Free-tailed Bat (Otomops martiensseni) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Large-eared Free-tailed Bat (Otomops martiensseni) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Côte d’Ivoire | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
There are no known studies on the likely effect of climate change of the ecology and distribution of the species. However, inferences can be made from ecological niche modelling studies (Ralph et al. 2015; Moir et al. 2020).
The species distribution appears to be mostly influenced by precipitation, as ecological niche models indicate that occurrence probabilities are markedly reduced in regions receiving <1000 mm of annual precipitation (Ralph et al. 2015). Such findings were corroborated by Moir et al. (2020), who showed precipitation of the driest month, as well as mean diurnal temperature range, and isothermality as important predictive bioclimatic factors in the distribution of the species in southern Africa. Changes in climate that affect annual precipitation, and that within the driest months, are therefore likely to result in a shift in distribution pattern within the assessment region.
Population information
This species occurs in isolated regions across a broad distributional range and is considered rare. For example, of 32 sites surveyed in Kruger National Park (KNP), it accounted for 0.74% of 11,655 echolocation call sequences and similarly comprised 0.16% of 1,284 call sequences in Mapungubwe National Parks (MNP) (Adams et al. 2015). However, past surveys have proved that this species can be locally common in certain areas (Long 1995; Taylor 1998; Fenton et al. 2002; Skinner & Chimimba 2005). For example, within the residential and agricultural areas around Durban, eThekwini Municipality, KwaZulu-Natal, it was once regularly observed (Fenton et al. 2002; Monadjem et al. 2010). However, there have been very few recent records of adult males from eThekwini Municipality (K. Richardson unpubl. data), and thus field surveys are required to ascertain current occupancy. In eThekwini, colonies consist of 30 or fewer individuals (Fenton et al. 2002). While colony size estimates are not available from Limpopo, we tentatively infer that that there are 10–30 mature individuals in each colony within the assessment region. Thus, based on 33 identified localities, this corresponds to a total mature population size of 330–990 animals. This could be an underestimate, based on undiscovered subpopulations, which is corroborated by the observation that this species is difficult to sample given that it forages at altitudes exceeding 600 m (Fenton & Griffin 1997), making trapping in mist-nets difficult. However, it could also be an overestimate, as past surveys in the eThekwini region have not been able to identify even 20 extant colonies. Further field data and research are needed to estimate population size more accurately.
In other parts of southern Africa, fewer than 10 specimens of this species have been collected (Monadjem et al. 2020). Globally, this species is suspected to be declining (but probably at a rate of less than 30% over ten years), owing to the probable continued loss of the known major East African populations, where O. martiensseni and O. harrisoni occur in sympatry (Patterson et al. 2018), while smaller subpopulations from southern Africa may be increasing (Mickleburgh et al. 2008).
Current population trend: Stable
Continuing decline in mature individuals? Unknown
Number of subpopulations: Unknown, but 33 colonies
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Suspected
All individuals in one subpopulation: Unknown
Number of mature individuals in largest subpopulation/colony: <1000
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
A phylogeographic study involving O. martiensseni was able to extend the distribution of the species into Kenya (Patterson et al. 2018). The study found no geographic structuring among sampling localities, with individuals from South Africa isolated to a clade interspersed with individuals from Rwanda, Kenya, Zimbabwe, Tanzania and the Ivory Coast. Given the genetic markers used, limited number of samples, and the vast majority of South African samples arising from KZN, a comprehensive understanding of the population genetic structure within the assessment region is not possible. However, it is likely that the species exists as a single metapopulation, connected at least to neighbouring Zimbabwe and possibly further north depending on its dispersal capabilities.
While the number of mature individuals has been estimated at ~1000 individuals within the assessment region, this cannot be translated to an effective population size given that the species likely exists as a metapopulation with neighbouring countries. To better gauge the species’ population genetic structure and diversity within the assessment region, a fine-scale nuclear genomic study is highly recommended.
Habitats and ecology
This species has been recorded from a range of habitats throughout Africa, from montane forest to savannah/woodland habitats in eastern Africa, up to altitudes of 2,000 m asl (Skinner & Chimimba 2005). Across their range, they are known to roost within caves, tree hollows or buildings (Fenton et al. 2002). Within the eThekwini region they will forage in areas of agriculture, such as areas of intensive sugar cane farming, and areas of urban development; and roost in urban and semi-urban areas, often utilising the roof rafters and inner brick surfaces of houses and other buildings (Fenton et al. 2002). Roost sites have not been located yet in Mapungubwe National Park, and Mbotyi and Manubi State Forests, but it seems plausible that the presence of human buildings and other infrastructure would provide roosting opportunities for colonies (Adams et al. 2015; Moir et al. 2020).
The Large-eared Giant Mastiff Bat is so named because they have extraordinarily large, rounded ears, which lie flat along the nose (Skinner & Chimimba 2005). This species has large, narrow wings, which allow for long-distance flight (Schoeman & Jacobs 2008), and although there is no direct evidence of migration, in East Africa, seasonal absence of the sister species O. harrisoni at important colony sites, indicates that migration may occur (Mutere 1973). Similarly, Adams et al. (2015) found this species to be more active in Kruger National Park (KNP) during the dry season than in the wet season, which supports the suggestion that foraging distances may be longer during the dry season. They are rapid fliers but are incapable of taking off from a flat surface, thus need to drop vertically from their roosting sites as they take off (Skinner & Chimimba 2005). Otomops spp. can perform unique flight manoeuvres involving sequences of slide-slips that alternate to the left and right when making steep descents into caves (Norberg & Rayner 1987). From faecal studies in Ethiopia the diet of O. harrisoni, was found to consist predominantly of Lepidoptera, as well as a small percentage of Hemiptera and Orthoptera (Rydell & Yalden 1997). The same holds true for O. martiensseni as it has a similar jaw structure to that of O. harrisoni and is designed for specialising in soft-bodied insect prey such as Lepidopterans (Freeman 1981; M.C. Schoeman unpublished data).
In the eThekwini area, this species occurs in colonies of fewer than 30 individuals, which usually consist of single males or females, male–female pairs or stable harems with a single male and 10 or more females with their young (Richardson & Taylor 1995; Taylor 1998; Taylor et al. 1999). However, in lava tunnels in East Africa, colonies of hundreds of individuals have been recorded (Kingdon 1974), although these may be attributable to the sister species O. harrisoni. In males, sexual maturity is reached after approximately one year, when they are evicted from the colony (Richardson & Taylor 1995). Studies have shown that this species may have an extended breeding season, from September to April (Taylor 1998).
Ecosystem and cultural services: As this species is insectivorous, it may contribute to controlling insect populations (Boyles et al. 2011; Kunz et al. 2011), in particular Lepidoptera (M.C. Schoeman, unpublished data). Bats belonging to the family Molossidae often prey on the insect species which destroy crops (Boyles et al. 2011; Kunz et al. 2011). Ensuring a healthy population of insectivorous bats can thus result in a decrease in the use of pesticides.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.5. Forest -> Forest – Subtropical/Tropical Dry | – | Suitable | – |
| 1.6. Forest -> Forest – Subtropical/Tropical Moist Lowland | – | Suitable | – |
| 1.9. Forest -> Forest – Subtropical/Tropical Moist Montane | – | Suitable | – |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Unknown | – |
| 14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land | – | Suitable | – |
| 14.2. Artificial/Terrestrial -> Artificial/Terrestrial – Pastureland | – | Unknown | – |
| 14.3. Artificial/Terrestrial -> Artificial/Terrestrial – Plantations | – | Suitable | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Suitable | – |
| 14.5. Artificial/Terrestrial -> Artificial/Terrestrial – Urban Areas | – | Suitable | – |
Life History
Generation Length: Unknown
Age at Maturity: Female or unspecified: Suspected to be the same as males, which is one year of age (see Richardson & Taylor 1995)
Age at Maturity: Male: One year (Richardson & Taylor 1995)
Size at Maturity (in cms): Female: Forearm length = 6.28 ±0.179 cm (Monadjem et al. 2020)
Size at Maturity (in cms): Male: Forearm length = 6.41 ±0.315 cm (Monadjem et al. 2020)
Longevity: Unknown
Average Reproductive Age: Unknown
Maximum Size (in cms): Female forearm length = 6.60 cm (Monadjem et al. 2020); Males forearm length = 6.77 cm (Monadjem et al. 2020)
Size at Birth (in cms): Unknown
Gestation Time: Three months reported for colonies from Kenya (Mutere 1973; although unsure if this data relates to O. martiensseni or the sympatric O, harrisoni
Reproductive Periodicity: In eThekwini, young have been recorded during the late spring to late autumn, however it is not clear whether females only once within each year (Taylor 200)
Average Annual Fecundity or Litter Size: Suspected to be one young per female (Taylor 2000)
Natural Mortality: Nocturnal avian predators including bat hawks and owls
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
There is no evidence to suggest that this species is traded or harvested within the assessment region.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
The leading threat to this species across its range appears to be roost disturbance. Major colonies of Otomops spp. in East Africa seem to have disappeared through disturbance of their cave habitats (Mickleburgh et al. 2008). Monitoring of subpopulations in KwaZulu-Natal over the past 10 years has shown that this species is not stable yet is able to make use of human-modified landscapes for roost sites. However, localised threats still include roost disturbance and possible indirect poisoning through the use of toxic timber treatments and persecution from ill-informed homeowners (Fenton et al. 2002). While threats to the Limpopo sand Eastern Cape subpopulations are not yet identified, roost disturbance and removal of large trees used as roost sites are plausible threats. Agricultural expansion and the use of pesticides may also impact the prey populations on which this species depends.
An emerging threat to this species may be wind farm development in KwaZulu-Natal and the expansion of wind energy facilities in the Eastern Cape to areas where the species occurs, due to the open-air foraging behaviour of, and large nightly distances covered by this species. The species is presently categories as “High-risk” to collision with wind turbines / barotrauma (MacEwan et al. 2020). When bats fly near to turbine blades, they either collide directly with the blade or they experience barotrauma which is tissue damage caused by rapid excessive changes in air pressure near turbine blades (Baerwald et al. 2008; Rydell et al. 2010).
Conservation
For African populations, this species is listed as Appendix II (2006) under the Convention on the Conservation of Migratory Species of Wild Animals (CMS). Within the assessment region, it has been recorded from both Kruger National Park (KNP) and Mapungubwe National Parks (MNP) in Limpopo (Adams et al. 2015, Parker and Bernard 2018), throughout the greater eThekwini region including protected areas (LR Richards, unpublished data), and Mbotyi and Manubi State Forests in the Eastern Cape province (Moir et al. 2020). It is protected by provincial ordinance in KwaZulu-Natal. An IUCN Species Action Plan has been developed for this species (Hutson et al. 2001). Continued monitoring of roost sites is important for the conservation of this species. There is an urgent need to reassess the status of all known roosts (and to locate additional localities) to ascertain numbers and status of colonies, so that key sites can be identified (Mickleburgh et al. 2008). Subsequently, disturbance of key localities should be restricted or managed to reduce the impact on colonies. Additionally, more research is necessary to better understand the natural history of this species and its relationship to habitat modification to develop a comprehensive and effective conservation plan (Adams et al. 2015).
Recommendations for land managers and practitioners:
- Reduce pesticide use in agricultural landscapes.
- Identification and protection of key roosting sites.
Research priorities:
- Further acoustic studies to determine the extent of the species distribution in the South African highveld.
- Systematic field surveys to determine distribution and population size and trends throughout the assessment region.
- Quantification of threats facing this species. This includes monitoring the impact of wind farming on Large-eared Giant Mastiff Bat populations within the Eastern Cape, KwaZulu-Natal and Mpumalanga provinces of South Africa and investigations into effective mitigation methods to reduce bat mortality around wind farms.
- Basic research into life history and ecology.
Encouraged citizen actions:
- Citizens can assist the conservation of the species by reporting sightings on virtual museum platforms (for example, iNaturalist and MammalMAP) and therefore contribute to an understanding of the species distribution.
- As the species emits audible vocalisations, citizens can use open-source software (e.g. Wildlife Acoustics SongMeter Touch) for recording echolocation calls. The recordings can be uploaded to iNaturalist.
Bibliography
Adams, R.A., Bonaccorso, F.J. and Winkelmann, J.R. 2015. Revised distribution for Otomops martiensseni (Chiroptera: Molossidae) in southern Africa. Global Ecology and Conservation 3: 707–714.
Baerwald, E.F., D’Amours, G.H., Klug, B.J. and Barclay, R.M. 2008. Barotrauma is a significant cause of bat fatalities at wind turbines. Current Biology 18: 695–696.
Brinkley, E.R., Weier, S.M., Parker, D.M. & Taylor. P.T. 2021. Three decades later in the northern Kruger National Park: multiple acoustic and capture surveys may underestimate the true local richness of bats based on historical collections. Hystrix, the Italian Journal of Mammalogy.
Boyles, J.G., Cryan, P.M., McCracken, G.F. and Kunz, T.H. 2011. Economic importance of bats in agriculture. Science 332: 41–42.
Cryan, P.M. and Barclay, R.M. 2009. Causes of bat fatalities at wind turbines: hypotheses and predictions. Journal of Mammalogy 90: 1330–1340.
Dorst, J. 1953. Notes on the genus and a description of a new species from Madagascar (Chiroptera, Molossidae). Mémoires de l’Institut Scientifique de Madagascar A8: 236–240.
Driver, A., Sink, K.J., Nel, J.N., Holness, S., van Niekerk, L., Daniels, F., Jonas, Z., Majiedt, P.A., Harris, L. and Maze, K. 2012. National Biodiversity Assessment 2011: An Assessment of South Africa’s Biodiversity and Ecosystems. Synthesis Report. South African National Biodiversity Institute and Department of Environmental Affairs, Pretoria, South Africa.
Fenton, M.B. et al. 2002. Researching little-known species: the African bat Otomops martiensseni (Chiroptera: Molossidae). Biodiversity & Conservation 11: 1583–1606.
Fenton, M.B. and Bell, G.P. 1981. Recognition of species of insectivorous bats by their echolocation calls. Journal of Mammalogy 62: 233–243.
Fenton, M.B. and Griffin, D.R. 1997. High-altitude pursuit of insects by echolocating bats. Journal of Mammalogy 78: 247–250.
Fenton, M.B., Jacobs, D.S., Richardson, E.J., Taylor, P.J. and White, W. 2004. Individual signatures in the frequency-modulated sweep calls of African large-eared, free-tailed bats Otomops martiensseni (Chiroptera: Molossidae). Journal of Zoology 262: 11–19.
Freeman, P.W. 1981. A multivariate study of the family Molossidae (Mammalia, Chiroptera): morphology, ecology, evolution. Fieldiana:. Zoology, new series 7: 1–173.
Hutson, A.M., Mickleburgh, S.P. and Racey, P.A. 2001. Microchiropteran Bats – Global Status Survey and Conservation Action Plan. IUCN/SSC Chiroptera Specialist Group, Gland, Switzerland and Cambridge, U.K.
Hutton, J.M. 1986. The status and distribution of bats in Zimbabwe. Cimbebasia 8: 219–236.
Kingdon, J. 1974. East African Mammals. Academic Press, London, UK.
Kunz, T.H., Braun de Torrez, E., Bauer, D., Lobova, T. and Fleming, T.H. 2011. Ecosystem services provided by bats. Annals of the New York Academy of Sciences 1223: 1–38.
Lamb, J.M. et al. 2006. Phylogeography of southern and northeastern African populations of Otomops martiensseni (Chiroptera: Molossidae). Durban Museum Novitates 31: 42–53.
Lamb, J.M.; Ralph, T.M.C.; Goodman, S.M.; Bogdanowicz, W.; Fahr, J.; Gajewska, M.; Bates, P.J.J.; Eger, J.; Benda, P. and Taylor, P.J. 2008. Phylogeography and predicted distribution of African-Arabian and Malagasy populations of giant mastiff bats, Otomops spp. (Chiroptera: Molossidae). Acta Chiropterologica 10: 21-40.
Long, J. K. 1995. Otomops martiensseni. Mammalian Species 493: 1-5.
MacEwan, K., Sowler, S., Aronson, J., and Lötter, C. 2020. South African Best Practice Guidelines for Pre-construction Monitoring of Bats at Wind Energy Facilities – ed 5. South African Bat Assessment Association.
Meester, J.A.J., Rautenbach, I.L., Dippenaar, N.J. and Baker, C.M. 1986. Classification of southern African mammals. Transvaal Museum Monographs 5: 1–359.
Mickleburgh, S., Hutson, A.M., Bergmans, W., Fahr, J. and Taylor, P.J. 2008. Otomops martiensseni.
Monadjem, A., Taylor, P.J., Cotterill, F.P.D. and Schoeman M.C. 2010. Bats of Southern and Central Africa: a biogeographic and taxonomic synthesis. University of Witwatersrand Press, Johannesburg.
Munyati, C. and Kabanda, T.A. 2009. Using multitemporal Landsat TM imagery to establish land use pressure induced trends in forest and woodland cover in sections of the Soutpansberg Mountains of Venda region, Limpopo Province, South Africa. Regional Environmental Change 9: 41–56.
Mutere, F. A. 1973. A comparative study of reproduction in two populations of the insectivorous bats, Otomops martiensseni, at latitudes 15°0’S and 2°30’S. Journal of Zoology (London) 171: 79-92.
Norberg, U.M. and Rayner, J.M. 1987. Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosophical Transactions of the Royal Society B: Biological Sciences 316: 335–427.
Parker, D.M. and Bernard, R.T. 2018. The use of acoustic detectors for assessing bat species richness and functional activity in a South African National Park. Mammalia 19;83(1):53-63.
Patterson, B.D., Webala, P.W., Bartonjo, M., Nziza, J., Dick, C.W. and Demos, T.C. 2018. On the taxonomic status and distribution of African species of Otomops (Chiroptera: Molossidae). PeerJ 24;6:e4864.
Ralph, T.M.C., Richards, L.R., Taylor, P.J., Napier, M.C. and Lamb, J.M. 2015. Revision of Afro-Malagasy Otomops (Chiroptera: Molossidae) with the description of a new Afro-Arabian species. Zootaxa 4057: 1– 49.
Richards, L.R.; Taylor, P.J.; Schoeman, M.C.; Goodman, S.M.; Van Daele, P.A.A.G. and Lamb, J.M. 2012. Cranial size and shape variation in Afrotropical Otomops (Mammalia: Chiroptera: Molossidae) testing species limits using a morphometric approach. Biological Journal of the Linnean Society 106(4): 910-925.
Richardson, E.J. and Taylor, P.J. 1995. New observations on the largeeared free-tailed bat Otomops martiensseni in Durban, South Africa. Durban Museum Novitates 20: 72–74.
Rydell, J., Bach, L., Dubourg-Savage, M-J., Green, M., Rodrigues, L. and Hedenström, A. 2010. Mortality of bats at wind turbines links to nocturnal insect migration? European Journal of Wildlife Research 56: 823–827.
Rydell, J. and Yalden, D.W. 1997. The diets of two high-flying bats from Africa. Journal of Zoology 242: 69–76.
Schoeman, M.C. and Jacobs, D.S. 2008. The relative influence of competition and prey defences on the phenotypic structure of insectivorous bat ensembles in southern Africa. PLoS one 3(11): e3715.
Simmons, N.B. 2005. Order Chiroptera. In: D.E. Wilson and D.M. Reeder (eds), Mammal Species of the World, pp. 312-529. The Johns Hopkins University Press, Baltimore, MD, USA.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Taylor, P.J., Cheney, C. and Sapsford, C. 1999. Roost habitat evaluation and distribution of bats (Chiroptera) in the Durban Metropolitan Region. Durban Museum Novitates 24: 62–71.
Taylor, P. 1998. The Smaller Mammals of KwaZulu-Natal. University of Natal Press, Pietermaritzburg, South Africa.
Taylor, P. 2000. Bats of Southern Africa: Guide to Biology, Identification, and Conservation. University of Natal Press, Pietermaritzburg.