

Grant’s Rock Rat
Micaelamys granti

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Micaelamys granti – (Wroughton, 1908)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – MURIDAE – Micaelamys – granti
Common Names: Grant’s Rock Rat, Grant’s Rock Mouse (English), Grant se Klipmuis (Afrikaans), Grant-Buschratte (German)
Synonyms: Aethomys granti (Wroughton, 1908)
Taxonomic Note: Although formerly included in the genus Aethomys, chromosomal, morphological and molecular evidence support the recognition of Micaelamys as a distinct genus (Baker et al. 1988, Russo 2003, Chimimba 2005, Lecompte et al. 2008). Micaelamys granti can be distinguished from M. namaquensis on morphological and chromosomal grounds (Visser and Robinson 1986, 1987; Chimimba et al. 1999). In M. namaquensis the tail is relatively longer, the ventral pelage is often pure white, and there are only three pairs of nipples; whereas, in M. granti, the ventral pelage is never pure white (usually grey or greyish), and there are five pairs of nipples (Monadjem et al. 2015).
Red List Status: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Russo, I.M.1
Reviewers: Kennerley, R.2
Institutions: 1Cardiff University,2IUCN SSC Small Mammal Specialist Group
Previous Assessors and Reviewers: Child, M.F., Avery, M., Palmer, G. & Birss, C.
Previous Contributors: Avenant, N., Baxter, R., MacFadyen, D., Monadjem, A., Taylor, P. & Wilson, B.
Assessment Rationale
This species is endemic to the Karoo regions of South Africa and is listed as Least Concern due to its wide distribution and because it exists in rocky habitat in a generally arid area that is unlikely to be transformed. Thus, there are no major threats that are suspected to be causing population decline. However, it is unknown whether this species can exist in agricultural landscapes and thus whether local declines are likely as climate change makes rocky habitats more suitable for agricultural expansion (for example, rooibos tea plantations in the Cedarberg). Field surveys are also required to collate localities where the species is currently present and thus enable the estimation of area of occupancy (AOO). If such data indicate a restricted AOO, this species could qualify for a threatened category given an inferred continuing decline in habitat. This species should be reassessed as new data become available.
Recommended citation: Russo IM. 2025. A conservation assessment of Micaelamys granti. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species is endemic to the Karoo regions of south-central South Africa, south of the Gariep River, where it occupies rocky outcrops in arid or semi-arid landscapes (Monadjem et al. 2015). It is sometimes sympatric with M. namaquensis (see Kok et al. 2012). It is confined to the southern and south-eastern areas of the Northern Cape Province, the northern and north-eastern areas of the Western Cape Province and the north-western regions of the Eastern Cape Province (Skinner and Chimimba 2005). Based primarily on museum records, it has an estimated extent of occurrence of 236,027 km². The species was sampled in 2000 in Loxton and Calvinia, Northern Cape (Russo, pers. comm.). There are very few recent (post-2000) records available. For example, no new records were discovered for this species from Barn Owl (Tyto alba) pellet analysis in either the Western or Northern Cape Provinces (Avery et al. 2005, Avery and Avery 2011), although it is perhaps not preyed upon extensively by Barn Owls. Recently, however, Kok et al. (2012) confirmed its presence in the Sneeuberg Mountain Complex (all sites higher than 1,700 m asl) of the Eastern Cape Province (sampling conducted between 2009 and 2010), where it occurred at low densities in the Sneeuberg Nature Reserve and Asante Sana Nature Reserve, but not the Mountain Zebra National Park. It is uncertain whether the overall absence of recent records is due to lack of current field surveys (low search effort) or a genuine loss in area of occupancy from anthropogenic transformation of habitat. Further vetting of museum specimens is also required.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
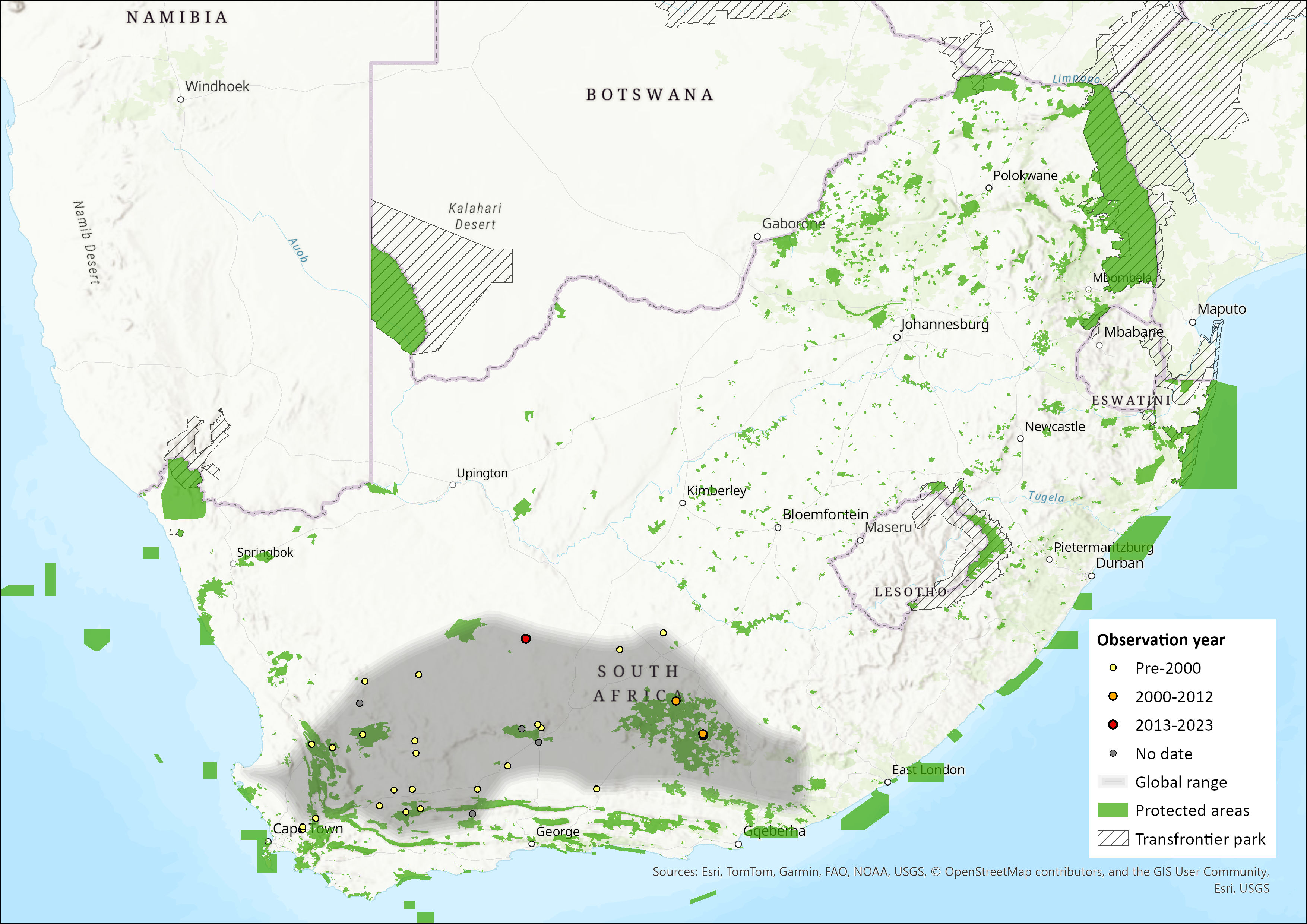
Figure 1. Distribution records for Grant’s Rock Rat (Micaelamys granti) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| South Africa | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Climate change makes rocky habitats more suitable for rooibos tea plantations such as in the Cederberg. It is however unknown whether this species can exist in agricultural habitats and thus whether local declines are likely to occur in the future.
Population Information
The abundance and population size are not known. However, the species is certainly rarer and far less abundant than M. namaquensis. For example, it comprised only 8.9% of total Micaelamys captures (N = 90 specimens) at three sites in the Sneeuberg Mountains, Eastern Cape Province (Kok et al. 2012). Its habitat currently does not appear to be extensively fragmented by human activities and thus the population is unlikely to be declining rapidly, but further field studies to assess occupancy and monitor the impacts of agricultural expansion into its habitats are needed to evaluate population trends. The lack of significant differences in cranial size across its geographical range suggests at least some connectivity between individuals and does not support any subspecies delineation (Chimimba et al. 1998).
Population Information
Current population trend: Stable (assumed)
Continuing decline in mature individuals: No
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: Naturally. Lives in high-altitude rocky habitats.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
A phylogenetic study (mitochondrial DNA cytochrome b sequences) showed that divergence time separating A. granti and A. namaquensis was estimated at 6.83 (range 6.41-7.28) MYR (million years) ago based on conventional substitution rate of 2% MYR (Russo 2003). In addition, a close evolutionary relationship between M. granti and M. namaquensis has been evident (Russo 2003).
While no population genetic studies have been undertaken on this species, its presumed continuous habitat and lack of morphological differences across its range suggest that the species could exist as a single metapopulation. From this, one of the two genetic indicators in the Convention of Biological Diversity’s Global Biodiversity Framework can be quantified – the complementary indicator- proportion of populations maintained with a species with 1 of 1 populations remaining, resulting in a value of 1.0 for this indicator.
Due to the lack of population estimates, the GBF’s headline indicator – proportion of populations with Ne greater than 500 – cannot be quantified.
It is recommended that a population genetic study be undertaken to get a better idea of the genetic structure and health/diversity within this species.
Habitats and ecology
This species occurs in shrubland and rocky areas on mountain slopes. It is not known if it can persist in disturbed or modified habitats, such as agricultural landscapes. It is likely its biology and ecology are similar to M. namaquensis.
Ecosystem and cultural services: Potential flagship species of the Karoo for biodiversity stewardship agreements.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 6. Rocky areas (eg. inland cliffs, mountain peaks) | – | Suitable | – |
Life History
| Generation Length | Justification | Data Quality |
| 1-2 | see Pacifici et al. 2013 | medium |
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species is not traded or utilised in any form.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
There are no known major threats as its habitat is largely intact and inaccessible. However, although, no threats were listed for this species in the previous assessment (Friedmann and Daly 2004), habitat loss from new forms of agricultural expansion may represent an emerging threat to this species. For example, the clearing of vegetation patches in rocky areas in the Cederberg region for rooibos tea plantations may lead to localised declines.
Habitat is stable with localised declines. The majority of the habitat will likely remain unchanged as it lives in rocky areas unsuitable for transformation. However, climate change is projected to make higher-altitude habitats more suitable for agriculture. For example, the suitability of upslope habitats for viticulture is projected to increase the footprint of winelands by 14% by 2050 (Hannah et al. 2013). Such trends in transformation of mid and upper slopes should be monitored
Conservation
We assume this species occurs in several protected areas within its range, for example, along the Cape Fold Mountain belt. However, its current presence has recently only been confirmed for Sneeuberg Nature Reserve and Asante Sana Nature Reserve in the Eastern Cape Province (Kok et al. 2012).
No specific conservation interventions are necessary, but it will benefit from protected area expansion to connect habitat and thus increase resilience to climate change and the effects of agricultural expansion onto higher elevations. Biodiversity stewardship schemes should also be pursued to achieve this, particularly if the landowner possesses pristine rocky outcrops and shrublands.
Recommendations for land managers and practitioners:
- Conserve/restore buffer strips of natural vegetation around rocky outcrops
Research priorities:
- Field surveys are needed to collate current distributional data and thus estimate area of occupancy
- There is no information on their habits, food or reproduction. Thus, basic ecological and biological information is required.
Encouraged citizen actions:
- Landowners and city planners can conserve natural vegetation around rocky outcrops.
Bibliography
Avery, D.M. and Avery, G. 2011. Micromammals in the Northern Cape Province of South Africa, past and present. African Natural History 7: 9-39.
Avery, D.M., Avery, G. and Palmer, N.G. 2005. Micromammalian distribution and abundance in the Western Cape Province, South Africa, as evidenced by Barn owls Tyto alba (Scopoli). Journal of Natural History 39: 2047–2071.
Baker, R.J., Qumsiyeh, M.B. and Rautenbach, I.L. 1988. Evidence for eight tandem and five centric fusions in the evolution of the karyotype of Aethomys namaquensis A. Smith (Rodentia: Muridae). Genetica 76: 161–169.
Chimimba, C.T. 2005. Phylogenetic relationships in the genus Aethomys (Rodentia: Muridae). African Zoology 40: 271–284.
Chimimba, C.T., Dippenaar, N.J. and Robinson, T.J. 1998. Geographic variation in Aethomys granti (Rodentia: Muridae) from southern Africa. Annals of the Transvaal Museum 36: 405-412.
Chimimba, C.T., Dippenaar, N.J. and Robinson, T.J. 1999. Morphometric and morphological delineation of southern African species of Aethomys (Rodentia: Muridae). Biological Journal of the Linnean Society 67: 501–527.
Friedmann, Y. and Daly, B. 2004. Red Data Book of the Mammals of South Africa: A Conservation Assessment. Conservation Breeding Specialist Group (SSC/IUCN) and Endangered Wildlife Trust, Parkview, South Africa.
Hannah, L., Roehrdanz, P.R., Ikegami, M., Shepard, A.V., Shaw, M.R., Tabor, G., Lu, Z., Marquet, P.A. and Hijmans, R.J. 2013. Climate change, wine, and conservation. Proceedings of the National Academy of Sciences of the United States of America 110: 6907-6912.
IUCN. 2017. The IUCN Red List of Threatened Species. Version 2017-2. Available at: www.iucnredlist.org. (Accessed: 14 September 2017).
Kok, A.D., Parker, D.M. and Barker, N.P. 2012. Life on high: the diversity of small mammals at high altitude in South Africa. Biodiversity and Conservation 21: 2823–2843.
Lecompte, E., Aplin, K., Denys, C., Catzeflis, F., Chades, M. and Chevret, P. 2008. Phylogeny and biogeography of African Murinae based on mitochondrial and nuclear gene sequences, with a new tribal classification of the subfamily. BMC evolutionary Biology 8: 199.
Monadjem, A., Taylor, P.J., Denys, C. and Cotterill, F.P.D. 2015. Rodents of Sub-Saharan Africa – a biogeographic and taxonomic synthesis. De Gruyter, Berlin/Munich/Boston.
Pacifici, M., Santini, L., Di Marco, M., Baisero, D., Francucci, L., Grottolo Marasini, G., Visconti, P. and Rondinini, C. 2013. Generation length for mammals. Nature Conservation 5: 87–94.
Russo, I-R. 2003. Molecular systematics of southern African Aethomys (Rodentia: Muridae). M.Sc. Thesis. University of Pretoria, Pretoria, South Africa.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Visser, D.S. and Robinson, T.J. 1986. Cytosystematics of the South African Aethomys (Rodentia: Muridae). South African Journal of Zoology 21: 264–268.
Visser, D.S. and Robinson, T.J. 1987. Systematic implications of spermatozoan and bacular morphology for the South African Aethomys. Mammalia 51: 447–454.