

Grant’s Golden Mole
Eremitalpa granti granti

2025 Red list status
Vulnerable
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Eremitalpa granti granti – (Broom, 1907)
ANIMALIA – CHORDATA – MAMMALIA – AFROSORICIDA – CHRYSOCHLORIDAE – Eremitalpa – granti
Common Names: Grant’s Golden Mole, Grant’s Desert Golden Mole (English), Grant se Gouemol, Woestynkruipmol (Afrikaans)
Synonyms: Chrysochloris granti Broom, 1907
Taxonomic Note:
Revised by Meester (1964). Two subspecies are recognised: E. g. granti (localised to South Africa) and E. g. namibensis (restricted to Namibia) based on differences in skull shape and pelage (Bronner and Jenkins 2005, Bronner 2013). Recent cytogenetic data (Gilbert et al. 2008) and ongoing phylogenetic analyses based on molecular, cytogenetic and morphological characters indicate that the subspecies are highly divergent and may represent cryptic species. A cryptic genetic lineage was recently detected in the vicinity of the Groen River mouth, near Garies, and this could represent a third subspecies or species (Mynhardt et al. 2024).
Red List Status: VU – Vulnerable, B1ab(iii)+2ab(iii) (IUCN version 3.1)
Assessment Information
Assessor: Mynhardt, S.1 & da Silva, J.2
Reviewer: Asher, R.J.3
Institutions: 1Endangered Wildlife Trust, 2South African National Biodiversity Institute, 3University of Cambridge
Previous Assessors & Reviewers: Maree, S. & Bronner, G.
Assessment Rationale
Eremitalpa granti granti is endemic to South Africa and is known from at least five locations along the coasts of the Western and Northern Cape provinces, with an estimated area of occupancy of 80 km2 (assuming a 16 km2 grid cell area), partially sufficient to warrant Endangered status under criteria B2ab(iii,iv). However, it is probably more widespread than current records suggest, as the northernmost (Port Nolloth) and southernmost (St Helena Bay) localities are 630 km apart on the continuous Namaqua Coastal Plain, and this subspecies is known to occur at least 20 km inland (for example, at Compagnies Drift near Lamberts Bay). Subopulations are probably not severely fragmented at present, and the extent of occurrence is likely to be > 12,000 km2. Although habitat loss and disturbance at multiple locations due to alluvial mineral mining of coastal dunes along the Northern Cape coastline (Kleinsee to Alexander Bay), the amount of available habitat remaining within the entire subspecies range is deemed to exceed the thresholds for Endangered listing under criterion B1b(iii). However, rapidly increasing mining activities in the area pose a significant threat to the remaining available habitat. The species is conserved in one protected area, so overall declines in area of occupancy, numbers of populations and population sizes are unlikely. However, listing of this subspecies as Vulnerable under criteria B1ab(iii) and/or B2ab(iii) is warranted given the estimates for area of occupancy and extent of occurrence above. Further field surveys are required to more accurately delimit range and occupancy.
Regional population effects: The Namibian subspecies is likely to be elevated to species status (Mynhardt et al. 2024), rendering the South African subspecies as an endemic species as the Orange River poses a barrier to dispersal. Thus, no rescue effect is possible.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recomended citations: Mynhardt S & da Silva JM. 2025. A conservation assessment of Eremitalpa granti granti. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Distribution
Geographic Range
The South African subspecies (E. granti granti) is confined to the west coast of southern Africa, from St Helena Bay (Western Cape Province, South Africa) northwards to Port Nolloth (and possibly as far north as Alexander Bay), and inland to Garies and the Biedouw Valley on the north-western aspect of the Cedarberg Mountains (Perrin & Fielden 1999; Bronner 2013; Taylor et al. 2018). The Orange River is likely to be the major biogeographical barrier that prevents gene flow and which has led to differentiation of E. g. granti and E. g. namibensis.
E. g. namibensis is confined to the west coast of southern Africa, from St. Helena Bay (Western Cape Province, South Africa) northwards to Walvis Bay (Namibia). E. g. granti occurs along the coast from south of St. Helena Bay to Port Nolloth (and possibly as far north as Alexander Bay), and inland to Garies and the Biedouw Valley on the north-western aspect of the Cedarberg Mountains. E. g. namibensis occurs in Namibia along the coast from the Orange River northwards to Walvis Bay, possibly as far as the Kuiseb River; and into the Namib Desert westwards as far as the ProNamib (Perrin & Fielden 1999, Bronner 2013, Taylor et al. 2018)
Collection efforts near Pt. Nolloth in the early 20th century (when, anecdotally, local children received rewards for finding specimens) evidently confused specimens of Eremitalpa granti with Cryptochloris wintoni, as jars labeled “Eremitalpa” accessioned in the Riksmuseet, Stockholm contained spirit-stored cadavers of both species. Eremitalpa granti and Cryptochloris wintoni therefore overlap in their ranges, but may exist in distinct microhabitats (Mynhardt et al. 2024) and are very divergent genetically and anatomically (Bronner et al. 2024).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
MAP
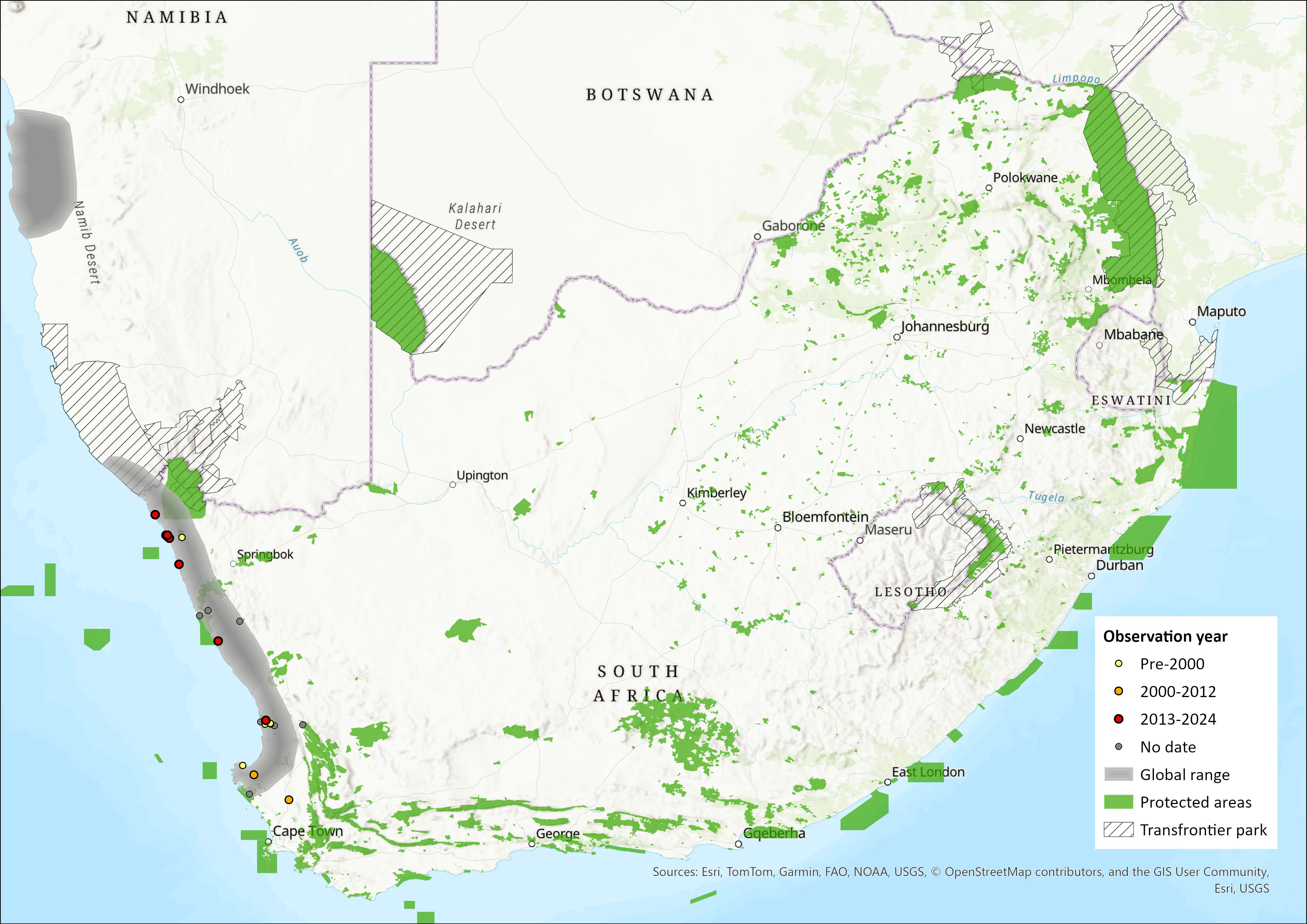 Figure 1. Distribution records for Grant’s Golden Mole (Eremitalpa granti granti) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for Grant’s Golden Mole (Eremitalpa granti granti) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| South Africa | Extant | Native | – | Resident |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Although the potential impacts of climate change on this species have not been directly assessed, climate change is expected to impact most golden mole species. Since these subterranean animals are restricted to soft soils for burrowing, and have very low vagility, their ability to migrate or shift their distribution ranges in response to climate change is very limited. Climate change is likely to impact golden moles through changes in vegetation type, soil moisture and prey availability, and coastal species additionally through reduced available habitat in developed/developing areas, as rising sea levels reduce the area of undeveloped coastal dune habitats between the coast and its associated developments.
Population information
Little is known about the population biology of the South African subspecies (E. g. granti). Numerous studies on E. g. namibensis in the Namib Desert indicate that densities are low (0.014–1.19 individual / ha) and home ranges large (3.1–12.3 ha) owing to the arid, energy-sparse conditions of their environment and widely dispersed prey availability. Although adults are solitary, spatial home range overlap is tolerated (Perrin & Fielden 1999).
Population Information
| Continuing decline in mature individuals? | Qualifier | Justification |
| Unknown | – | – |
Current population trend: Unknown
Extreme fluctuations in the number of subpopulations: (Not specified)
Continuing decline in number of subpopulations: (Not specified)
All individuals in one subpopulation: No
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Unknown
Severely fragmented: Unknown, may be fragmented by diamond mining activities and poor dispersal capacity.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
A recent molecular study revealed genetic differentiation between the two subspecies (and possibly a third) and has suggested the Orange River is likely to be the major biogeographical barrier preventing gene flow between them (Mynhardt et al. 2024). This could have E. g. namibensis (which is confined to the area north of the Orange river Orange River to Walvis Bay, possibly as far as the Kuiseb River; and into the Namib Desert westwards as far as the ProNamib (Perrin & Fielden 1999, Bronner 2013, Taylor et al. 2018)) elevated to full species status, resulting in the species being a South African endemic, occuring along the west coast from south of St. Helena Bay to Port Nolloth (and possibly as far north as Alexander Bay), and inland to Garies and the Biedouw Valley on the north-western aspect of the Cedarberg Mountains.
No estimate of effective population size is possible due to limited information.
Habitats and ecology
E. granti are limited to the Strandveld and Succulent Karoo biomes of South Africa. They prefer soft sands of coastal dune crests but are also present in inter-dune swales with quite dense vegetation as long as sand is not too consolidated. Areas containing scattered clumps of the Dune Grass (Aristida sabulicola), Ostrich Grass (Cladoraphis spinosa) and Long Bushman Grass (Stipagrostis ciliata), are the preferred habitats for this subspecies. The young are thought to be born in tunnels constructed by adults, but they lack a proper burrow system. Resting sites are usually under vegetation. It is a nocturnal surface forager that specialises on termites, but also consumes other invertebrates and small vertebrates (legless lizards, web-footed geckos and sand-burrowing skinks). It is nocturnal during hot summer months, with greater diurnal activity observed during winter (Fielden et al. 1990a, 1990b, 1992; Rathbun & Rathbun 2007). It has occasionally been recorded from arable land and rural gardens (for example, near Lamberts Bay).
Ecosystem and cultural services: This subspecies is not known to provide any specific ecosystem services, but this may simply reflect the paucity of information available for this poorly known subspecies. Eremitalpa granti has, however, commonly been recorded as a prey species for Barn Owls (Tyto alba). Additional predators such as Pied Crows (Corvus alba), Pale Chanting Goshawks (Melierax canorus), Spotted Eagle Owls (Bubo africanus), Striped Polecats (Ictonyx striatus) and Black-backed Jackals (Lupullela mesomelas) have also been documented (Skinner & Chimimba 2005).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 3.4. Shrubland -> Shrubland – Temperate | Resident | Suitable | No |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | Resident | Suitable | No |
| 3.8. Shrubland -> Shrubland – Mediterranean-type Shrubby Vegetation | Resident | Suitable | Yes |
| 8.3. Desert -> Desert – Cold | Resident | Suitable | Yes |
| 14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land | Resident | Suitable | No |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | Resident | Suitable | No |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: 7-9
Size at Maturity (in cms): Male: 7-9
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: Births recorded in October-November (austral spring)
Average Annual Fecundity or Litter Size: Litter size 1-2.
Natural Mortality: (Not specified)
Breeding Strategy
Does the species lay eggs? No
Does the species give birth to live young? Yes
Does the species exhibit parthenogenesis? No
Does the species have a free-living larval stage? No
Does the species require water for breeding? No
Movement Patterns
Movement Patterns: Not a Migrant
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This subspecies is not known to be utilised or traded in any form.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
Alluvial diamond mining in coastal areas in Northern Cape (Kleinsee and Port Nolloth) results in marked transformation and fragmentation of habitats at some locations due to the removal of topsoil (Smithers 1986). Rapidly increasing mining activities (evidenced by current mining and prospecting activities, as well as numerous mining applications) in the area pose a significant threat to the remaining available habitat. However, much of the range of this subspecies coincides with coastal desert where human influence on habitats is not substantial, so the overall population is probably not in decline. Coastal tourism developments along the south-western coast of South Africa (St Helena Bay to Lambert’s Bay), and agricultural practices on the Namaqualand coastal plain (and associated inland valleys) have resulted in some habitat transformation, but threats are considered minor as this subspecies can survive in mildly transformed habitats. Although rehabilitation of mined areas may be partially mitigating habitat loss, these localised impacts, together with the few known populations and limited EOO and AOO of the subspecies, are sufficient for threatened listing.
Habitat trend: Declining in area and quality. For example, rural and urban settlement expansion in the Northern Cape has increased by 9% and 15% respectively between 2000 and 2013 (GeoTerraImage 2015). Similarly, in the Western Cape, Pence (2014) calculated that between 2006 and 2011, 536 km2 of land was converted to agriculture (107 km2 / year, which equates to 0.08% of the surface area of the province per year).
Conservation
This subspecies is protected in the Namaqua National Park in South Africa, as well as some smaller privately owned conservation areas. Although rapidly increasing mining activities in the area pose a significant threat to the remaining available habitat, no specific conservation interventions are necessary at present. Although much of its range coincides with coastal desert where human influences on habitats are not substantial (including coastal tourism development and agricultural practices on the Namaqualand coastal plains), it can survive in mildly transformed habitats.
Recommendations for land managers and practitioners:
- Field surveys to locate additional subpopulations.
- Assessment of population viability within rehabilitated mining areas.
- Assessment of connectivity among subpopulations.
Research priorities:
- Ecological requirements and niche tolerances.
- Systematic surveying to accurately delimit distribution of both subspecies.
- Population genetic analysis including representatives of all known subpopulations representative of entire distribution range to identify and quantify population substructure and levels of historic and current gene flow.
- Studies assessing subpopulation trends and the severity of threats outside of protected areas.
- Research into population recoveries in rehabilitated mined areas.
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Deposit any dead specimens found in a state or provincial museum, together with information on the date and site where found.
- Create indigenous vegetation gardens.
Bibliography
Bronner, G.N. 2013. Eremitalpa granti. In: J. Kingdon, D. Happold, T. Butynski, M. Hoffmann, M. Happold and J. Kalina (eds), Mammals of Africa, Volume I: Introductory Chapters and Afrotheria, pp. 253-254. Bloomsbury Publishing, London.
Fielden, L.J., Hickman, G.C. and Perrin, M.R. 1992. Locomotory activity in the Namib Desert golden mole Eremitalpa granti namibensis (Chrysochloridae). Journal of Zoology (London) 226: 329–344.
Fielden, L.J., Perrin, M.R. and Hickman, G.C. 1990. Feeding ecology and foraging behaviour of the Namib Desert golden mole, Eremitalpa granti namibensis (Chrysochloridae). Journal of Zoology, London 220: 267–289.
Fielden, L.J., Waggoner, J.P., Perrin, M.R. and Hickman, G.C. 1990b. Thermolegulation in the Namib Desert golden mole, Eremitalpa granti namibensis (Chrysochloridae). Journal of Arid Environments 18: 221-237.
GeoTerraImage. 2015. Quantifying settlement and built-up land use change in South Africa. Pretoria.
Gilbert, C., Maree S. and Robinson T.J. 2008. Chromosomal evolution and distribution of telomeric repeats in golden moles (Chrysochloridae, Mammalia). . Cytogenetic and Genome Research 121(2): 110-119.
Meester J.A.J. 1964. Revision of the Chrysochloridae. I. The desert golden mole Eremitalpa Roberts. Scientific Papers of the Namib Desert Research Station 26: 1–7.
Mynhardt, S., Matthew, E., Le Roux, J.P., Little, I., Bloomer, P. and Theron, C., 2024. Environmental DNA from soil reveals the presence of a “lost” Afrotherian species. Biodiversity and Conservation 33(1), pp.31-50.
Perrin, M.R. and Fielden, L.J. 1999. Eremitalpa granti. Mammalian Species 629: 1-4.
Rathbun, G.B and Rathbun, C.D. 2007. Habitat use by radio-tagged Namib Desert golden moles (Eremitalpa granti namibensis). African Journal of Ecology 45: 196-201.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Smithers, R.H.N. 1986. South African Red Data Book – Terrestrial Mammals. South African National Scientific Programmes Report 125: 1–216.
Taylor, W.A., Mynhardt, S. and Maree, S. 2018. Family Chrysochloridae. In: Handbook of the Mammals of the World – Volume 8 Insectivores, Sloths and Colugos. DE Wilson & RA Mittermeier (eds). Lynx Edicions. ISBN-13: 978-84-16728-08-4.