

Common Mole Rat
Cryptomys spp.

2025 Red list status
Cryptomys h. hottentotus, Cryptomys h. mahali, Cryptomys h. natalensis, Cryptomys h. pretoriae

2025 Red List Status
Cryptomys h. nimrodi,
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Cryptomys spp. — Common Mole-rats
ANIMALIA — CHORDATA-MAMMALIA — RODENTIA — BATHYERGIDAE — Cryptomys
Common Names: All species: Common Mole-rat, African Mole-rat, Common Blesmol, Mole-rat (English), Vaalmol, Grysmol (Afrikaans); Cryptomys hottentotus: Hottentot Mole-rat (English); Cryptomys h. mahali: Mahali’s Mole-rat (English); Cryptomys h. natalensis: Natal Mole-rat; Cryptomys h. pretoriae: Highveld Mole-rat (English), Crytpomys h. nimrodi (Matabeleland mole-rat) (English).
Synonyms: Cryptomys hottentotus: abberans, albus, bigalkei, caecutiens, cradockensis, exenticus, holosericius, jamesoni, jolangi, kopmotiensis, langi, lugwigii, melanoticus, montanus, nemo, orangiae, rufulus, stellatus, talpoides, transvaalensis, valschensis, vandami, vrybergensis, zimibitiensis; Cryptomys mahali: amatus; Cryptomys natalensis: zuluensis
Taxonomic Note: The genus Cryptomys previously included all species now listed under the genus Fukomys. These two genera can be distinguished morphologically from one another but are considered highly divergent from one another on a molecular level (Faulkes et al. 2004, Ingram et al. 2004). Faulkes et al. (2004) lists five subspecies belonging to the genus Cryptomys, all considered endemic or near endemic to the assessment region (Bennett 2013):
- Common Mole-rat, C. h. hottentotus (tentatively Northern Cape, Western Cape, Eastern Cape).
- Natal Mole-rat, C. h. natalensis (tentatively KwaZulu-Natal, Eastern Cape, Mpumalanga and Limpopo provinces; Eswatini and Lesotho).
- Mahali Mole-rat, C. h. mahali (tentatively Gauteng, North West, Free State, and Northern Cape provinces).
- Highveld Mole-rat, C. h. pretoriae (tentatively Gauteng, North West Province).
- Matabeleland mole-rat C. h. nimrodi (tentatively Limpopo and southern Zimbabwe).
These five subspecies were previously grouped under a single species C. hottentotus, but were found to reveal significant molecular and differences worthy of reclassification (Faulkes et al. 2004). In general, they are considered allopatric, however the specific limits of each range are yet to be defined (Faulkes et al. 2004). These subspecies can currently only be distinguished from each other through molecular techniques. Currently there is no formal elevation of the subspecies to specific level (Bennett pers comm).
Red List Status:
- Cryptomys h. hottentotus, LC — Least Concern
- Cryptomys h. mahali, LC — Least Concern
- Cryptomys h. natalensis, LC — Least Concern
- Cryptomys h. pretoriae, LC— Least Concern
- Cryptomys h. nimrodi, DD — Data Deficient (IUCN version 3.1)
The genus Cryptomys shows the broadest geographical distribution of all African Mole-rats, and the various patterns of speciation within this genus are expected to have resulted from environmental changes during the Miocene (Faulkes et al. 2004).
Red List Assessment
Assessment Information
Assessors: Bennett, N1., Finn, K1. & da Silva, J2.
Reviewer: Patel, T3.
Institutions: 1University of Pretoria, 2South Africa, South African National Biodiversity Institute, Endangered Wildlife Trust3
Previous Assessors & Reviewers: Bennett, N.C., Jarvis, J., Visser, J. & Maree, S.
Previous Contributors: Wilson, B., Palmer, G., MacFadyen, D., Avery, M., Child, M.F., Cohen, L., Relton, C., Avenant, N., Baxter, R., Monadjem, A. & Taylor, P.
Assessment Rationale: The Common Mole-rat, C. h. hottentotus, Natal Mole-rat, C. h. natalensis and Mahali Mole-rat, C. h. mahali are listed as Least Concern because they are widely distributed throughout the assessment region, are reasonably common and adapt well to habitats modified by humans, such as agricultural areas and rural gardens. The Highveld Mole-rat, Cryptomys h. pretoriae, is also listed as Least concern, even though it has a restricted distribution to the grassland of Pretoria and Johannesburg suburbs. Currently, there are no major recognised threats that could result in significant population decline. The Matabeleland mole-rat, Cryptomys h. nimrodi is listed as Data Deficient, as it is only recorded in the northern Limpopo.
Regional population effects: Due to disjunctions in the distribution, dispersal is impossible for C. h. hottentotus and C. pretoriae. The dispersal of C. h. mahali and C. h. natalensis is currently unknown.
Red List Index: Cryptomys h. mahali — changed from DD to LC; Cryptomys h. nimrodi new assessment (DD)
Recommended citation: Bennett N, Finn K & da Silva JM. 2025. A conservation assessment of Cryptomys hottentotus pretoriae. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Distribution
Geographic Range
The Common Mole-rat, C. h. hottentotus, occurs throughout the Western Cape from the south-western limits of Cape Town and extends northwards the Northern Cape, Steinkopf region. It is expected to be the only Cryptomys subspecies within its range; however, the exact northern and eastern limits of its distribution remain undefined, due to confusion with other subspecies species in this genus (Bennett and Faulkes pers comm.). Mahali’s Mole-rat, C. h. mahali, is known from north of the Magaliesberg Ridge from near Patryshoek (Gauteng province) and Brits (North West) (Faulkes et al 2004; Bennett, Hart, Finn, unpubl. data). This species has also been identified in Van Zyls Rus (Northern Cape), Bloemhof (North West Province), and Bloemfontein (North West) (N.C. Bennett unpubl. data). Further research is required in order to establish the precise limits of the species’ range. The Natal Mole-rat, C. h. natalensis, is present throughout KwaZulu-Natal, Mpumalanga, Lesotho and Eswatini. Its range may extend into the Kruger National Park and extreme southern regions of Mozambique. C. h. natalensis may overlap in its range with C. h. pretoriae in Mpumalanga. The precise boundaries of its range are unresolved, and it is possible the point localities in the northeastern regions of South Africa (northern Kruger National Park) may belong to C. h. nimrodi rather than C. h. natalensis (Monadjem et al. 2015). The Highveld Mole-rat, C. h. pretoriae, occurs in Gauteng (Bennett 2013; Its eastern limit is likely to be the Mpumalanga Drakensberg escarpment, which is the western limit of C. h. natalensis (Faulkes et al. 2004). The Matabeleland Mole-rat, C. h. nimrodi is found in the northern Limpopo Province close to the Limpopo River in South Africa. The southern limit of the species range is Limpopo south of the Limpopo River and extends northwards to Bulawayo, Zimbabwe, but the eastern limit of its range is yet to be determined. For all species, further molecular research and vetting of museum records is required to more accurately delineate range distributions.
Elevation / Depth / Depth Zones
- Elevation Lower Limit: (Not specified)
- Elevation Upper Limit: (Not specified)
- Depth Lower Limit: (Not specified)
- Depth Upper Limit: (Not specified)
- Depth Zone: (Not specified)
Map
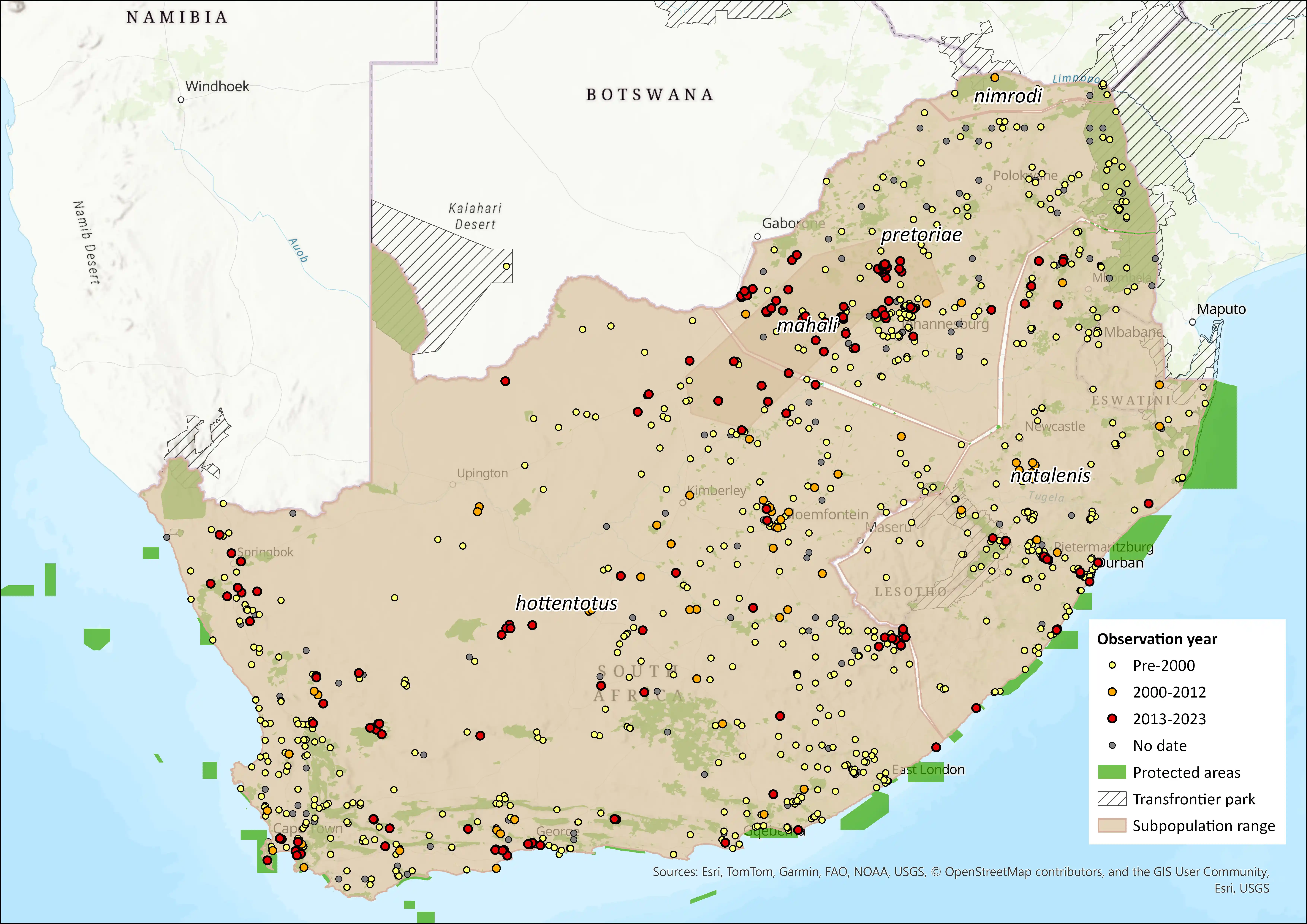 Figure 1. Distribution records for the Common Mole-rat (Cryptomys spp.) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Figure 1. Distribution records for the Common Mole-rat (Cryptomys spp.) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Countries of Occurrence
| Country | Presence | Origin |
| Botswana | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Absent | – |
| C. pretoriae | Absent | – |
| Lesotho | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Extant | Native |
| C. pretoriae | Absent | – |
| Mozambique | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Extant | Native |
| C. pretoriae | Absent | – |
| Namibia | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Absent | – |
| C. pretoriae | Absent | – |
| South Africa | ||
| C. hottentotus | Extant | Native |
| C. mahali | Extant | Native |
| C. natalensis | Extant | Native |
| C. pretoriae | Extant | Native |
| C. nimrodi | Extant | Native |
| Eswatini | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Extant | Native |
| C. pretoriae | Presence uncertain | Native |
| Zimbabwe | ||
| C. hottentotus | Absent | – |
| C. mahali | Absent | – |
| C. natalensis | Absent | Native |
| C. pretoriae | Absent |
–
|
| C. nimrodi | Extant |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Climate change may threaten to reduce suitable habitat for these species in the future; however, it is currently not a threat to these species, especially because they can survive in transformed habitats. Further research is needed on climate change and Cryptomys species.
Population information
The population size of this genus varies depending on the subspecies. The Mahali Mole-rat has a density of 5.5 animals/hectare; the Natal Mole-rat 4.9/hectare, the Highveld Mole-rat 2.2/hectare and the Common Mole-rat 5/hectare (Suess et al. 2024, Spinks et al, 2000).
However, it is widespread and abundant across the assessment region. Presumably the population is limited by soil requirements. In prime habitats the population density of C. h. hottentotus is known to surpass 150 individuals/km² (A.C. Spinks, J.U.M. Jarvis & N.C. Bennett unpubl. data). The populations are not predicted to be declining due to their broad distribution and their ability to survive successfully within transformed habitats.
Current population trend: Stable
Continuing decline in mature individuals? Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individual in one subpopulation:
Number of individuals in population: Many
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: No
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years:
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years:
Probability of extinction in the wild within 100 years:
Population genetics
While population genetic studies have not been conducted on each of the identified subspecies, a recent study looked at the structure and diversity of C. hottentotus hottentotus (Merchant et al. 2024) and found this subspecies to be divided into two metapopulations separated by an aridity gradient with a mesic and arid/semi-arid population identified. Moreover, the studies found greater genetic diversity and gene flow in the arid metapopulation, suggesting greater connectivity and interactions between colonies in arid regions in comparison to mesic ones. Given these findings, it is possible additional substructure and varying levels of diversity exist in the other subspecies and therefore they should be investigated further. Assuming no populations have gone extinct and that the other subspecies exist as a single metapopulation, the Convention on Biological Diversity’s complementary genetic indicator — proportion of populations maintained (PM) — would receive a value of 1.0 (6/6 populations remain). Based on the available density estimates, it would seem fair to estimate each population’s general size to be at least 5,000 individuals. Applying a conversion ratio of 0.1-0.3 effective population size to census size (Ne/Nc) we would estimate the Ne of each population to exceed 500, suggesting they are genetically stable and healthy. However, given their social structure a DNA-based calculation of Ne is highly recommended to obtain more accurate estimates. This would be possible for C. h. hottentotus given that a RadSeq dataset already exists.
Habitats and ecology
Generally, the subterranean Cryptomys subspecies occur across a wide range of soil types from fine to medium grained clays and loams. They are often located within human-modified environments, such as lawns, golf courses and gardens (particularly in the presence of vegetable gardens). They specialise in bulbs, corms and tubers, especially those of Albuca, Lachenalia, Ornithogalum (Hyacinthaceae), Romulea, Micranthus, Homeria (Iridaceae) and Oxalis (Oxalidaceae) (Bennett 2013).
The Common Mole-rat, C. h. hottentotus, is social. Colonies consist of a single breeding pair. Mean colony size is 5.1, but colonies can attain a size of 15 animals in their mesic distribution or a maximum of 11 in their arid distribution (Bennett pers obs.). It is a seasonal breeder producing 1 to 2 litters per annum with a mean litter size of 2.3 in the mesic habitat and 2.6 in the arid regions. The mean body mass of males is 83±22g and in females 58±14g. The breeding season is from September to November. Mean colony biomass ranges from 386g in mesic habitats to 287g in arid habitats. Colony density is 1.8 colonies/hectare in arid habitats and 16 colonies/hectare in mesic habitats. (Spinks et al. 2000).
Mahali’s Mole-rat, C.h. mahali, is found in more arid habitats when compared to C. h. pretoriae, and is typically located in clays and sandy soils in scrubland habitats. Colonies comprise of a single breeding pair and a number of non-breeders of both sexes. Mean colony size is 7.7, but colonies can attain a maximum size of 24. It is an a-seasonal breeder producing 2 to 3 litters per annum with a mean litter size of 1.9±0.8. The mean body mass of males is 104±28 and females 76±12. The breeding period is from January to December. The mean biomass of the colony is 507g. Colony density is 1.5 colonies per hectare (Suess et al. 2024).
The Natal Mole-rat, C. h. natalensis, is found in clay and loam soil where it feeds upon corms, bulbs and rhizomes of grasses. It is found predominantly in grasslands (occasionally including montane grasslands), and occurs sympatrically with the Hottentot Golden Mole, Amblysomus hottentotus, which can frequently be found sharing the same tunnel. This species is colonial, with up to 20 individuals in a single colony. The colony has a single breeding pair and subordinates of both sexes. Mean colony size is 6.6, but a colony of 20 animals has been captured. It is an aseasonal breeder producing 2 to 3 litters per annum with a mean litter size of 1.4±0.6. The mean body mass of males is 93.1±22g and females 80.3±14g. The breeding season is from January to December. The mean biomass of a colony is 535g. Colony density is 1.2 colonies per hectare (Suess et al., 2024).
The Highveld Mole-rat, C. h. pretoriae, is found in an array of soils from clay and sandy through to loams. In contrast to C. h. hottentotus (which tends to avoid stony soils), this species will venture into stony soils. They are predominantly a grassland species, feeding on grass rhizomes and any associated geophytes in the area. Again, this species is colonial, consisting of group sizes up to 15 individuals (Moolman et al. 1998). This species thrives in Highveld grasslands, but is also located within Burkhia scrubland. Similar to other Cryptomys species, they are able to survive in modified habitats and in particular gardens and golf courses but are also found on vacant plots and cultivated lands. The colony has a single breeding pair and subordinates of both sexes. Mean colony size 4.0, but 15 animals have been recorded. It is a seasonal breeder producing 1 or occasionally 2 litters per annum with a mean litter size of 1.3±0.5 pups. The mean body mass of males is 120±35g and females 98±20g. The breeding season is from July to November. The mean biomass of a colony is 410g. Colony density is 1.2 colonies per hectare (Suess et al. 2024).
Ecosystem and cultural services: Generally, Mole-rats are good ecosystem engineers, as they enrich the humus content of the soil, aerate the soil, assist with drainage systems and enhance plant species diversity (Hagenah & Bennett 2013). Additionally, they create refuges for other species to use to escape fire. Common Mole-rats also eat geophytes that contain cardiac glycosides (for example, Ornithogalum spp., Homeria spp., Morea spp.), which are toxic to livestock (Bennett 2013). They are eaten by a wide range of predators (Bennett 2013).
IUCN Habitats Classification Scheme
Life History
Age at maturity: female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
Table 1: Use and trade summary for the Cryptomus spp.
|
Category |
Applicable? |
Rationale |
Proportion of total harvest |
Trend |
|
Subsistence use |
Yes |
Bushmeat |
Unknown |
Stable |
|
Commercial use |
No |
– |
– |
– |
|
Harvest from wild population |
Yes |
Bushmeat |
Unknown |
Stable |
|
Harvest from ranched population |
No |
– |
– |
– |
|
Harvest from captive population |
No |
– |
– |
– |
Threats
No major threats have been identified for these species. However, they are occasionally persecuted as an agricultural pest, and homeowners complain that C. h. hottentotus and C. h. pretoriae can become nuisances in their gardens (Power 2014), and they are occasionally exterminated on golf courses.
Current habitat trend: Stable
Conservation
Cryptomys subspecies are located in numerous protected areas throughout the assessment region. For example, Bloemhof Nature Reserve, Kruger National Park and Nylesvley Nature Reserve, which are key protected areas for C. mahali, C.h. natalensis or nimrodi respectively. As a result, no specific interventions are necessary for these species. However, further research is necessary to delineate the population distribution, trends and abundance of these species.
- Recommendations: Systematic surveys needed to gather information on population size, trends and distribution.
- Research priorities: Accurate species distribution mapping is necessary through molecular research museum record vetting.
Encouraged citizen actions: Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas. However, subspecific level identification will require molecular analyses.
Bibliography
Bennett NC. 2013. Cryptomys hottentotus Common Mole-rat (Hottentot Mole-rat). Pages 655-658 in Happold DCD, editor. Mammals of Africa. Volume III: Rodents, Hares and Rabbits. Bloomsbury Publishing, London, UK.
Faulkes CG, Verheyen E, Verheyen W, Jarvis JUM, Bennett NC. 2004. Phylogeographical patterns of genetic divergence and speciation in African mole-rats (Family: Bathyergidae). Molecular Ecology 13: 613-629.
Hagenah N, Bennett NC. 2013. Mole rats act as ecosystem engineers within a biodiversity hotspot, the Cape Fynbos. Journal of Zoology 289:19-26.
Merchant, H.N., Ivanova, A., Hart, D.W., Garcia, C., Bennett, N.C., Portugal, S.J. and Faulkes, C.G. 2024. Patterns of Genetic Diversity and Gene Flow Associated With an Aridity Gradient in Populations of Common Mole-rats, Cryptomys hottentotus hottentotus, Genome Biology and Evolution, 16(7). evae144, https://doi.org/10.1093/gbe/evae144
Moolman M, Bennett NC, Schoeman AS. 1998. The social structure and dominance hierarchy of the highveld mole-rat Cryptomys hottentotus pretoriae (Rodentia: Bathyergidae). Journal of Zoology 246:193-201.
Power R.J. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Rautenbach IL. 1982. Mammals of the Transvaal. No. 1, Ecoplan Monograph. Pretoria, South Africa.
Spinks, A.C. Bennett, N.C., and Jarvis, J.U.M. (2000). A comparison of the ecology of two populations of the common mole-rat, Cryptomys hottentotus hottentotus: the effect of aridity on food, foraging and body mass. Oecologia 125(3): 341-349.
Spinks, A.C., Jarvis, J.U.M. and Bennett, N.C. (2000). Comparative patterns of philopatry and dispersal in two mole-rat populations: implications for the evolution of mole-rat sociality. Journal of Animal Ecology 69(2): 224-234.
Suess, T., Finn, K.T., Janse van Vuuren, A.K., Hart, D.W. and Bennett, N.C. (2024). A comparison of the population ecology of 4 Cryptomys hottentotus subspecies. Journal of Mammalogy 105: 26-39.