

Cape Mole-rat
Georychus capensis

2025 Red list status

2025 Red list status
2025 Red list status
Regional Population Trend
Unknown
Change compared
to 2016
Overview
Georychus capensis – (Pallas, 1778)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – BATHYERGIDAE – Georychus – capensis
Common Names: Cape Mole Rat, Cape Blesmol (English), Kaapse Blesmol (Afrikaans)
Synonyms: Fossor leucops Lichtenstein, 1844; Georychus capensis ssp. canescens Thomas & Schwann, 1906; Georychus yatesi Roberts, 1913; Mus buffoni Cuvier, 1834
Taxonomic Note: The subpopulations from KwaZulu-Natal (Honeycutt et al. 1987, Nevo et al. 1987, J.H. Visser unpubl. data) and Mpumalanga (C.G. Faulkes and N.C. Bennett unpubl. data; J.H. Visser unpubl. data) are genetically distinct from each other, and ongoing molecular research is uncovering at least two lineages in the Western Cape (J.H. Visser unpubl. data). The KwaZulu-Natal (Honeycutt et al. 1987, Nevo et al. 1987, J.H. Visser unpubl. data) and Mpumalanga (J.H. Visser unpubl. data) populations are considered worthy of species status, and other lineages may constitute species too. However, further research and taxonomic resolution for all subpopulations is needed.
Red List Status:LC – Least Concern, (IUCN version 3.1)
KwaZulu-Natal subpopulation – DD, Data Deficient
Mpumalanga subpopulation – DD, Data Deficient
Assessment Information
Assessors: Bennett, N.1 & da Silva, J.M.2
Reviewer: Patel, T.3
Institutions:1University of Pretoria, South Africa,2South African National Biodiversity Institute, 3Endangered Wildlife Trust
Previous Assessors & Reviewers: Bennett, N.C., Jarvis, J., Visser, J. & Maree, S.
Previous Contributors: Wilson, B., Palmer, G., MacFadyen, D., Avery, M., Child, M.F., Cohen, L., Relton, C., Avenant, N., Baxter, R., Monadjem, A. & Taylor, P.
Assessment Rationale
As a relatively common species, the Cape Mole-rat is currently listed as Least Concern. It is widely distributed across the south-western regions of South Africa and along the southern coastline into the Eastern Cape and can occupy human-modified landscapes although they require certain ecological variables to be met. Additionally, separate, disjunct subpopulations occurring in isolated localities in KwaZulu-Natal and Mpumalanga have been identified but are rarely recorded. There is no current data on population size or trend, and it is uncertain whether habitat loss and degradation is a threat to the species. Climate change may threaten the fragmented subpopulations by reducing suitable habitat in the future. Further research on the taxonomic and conservation status of these subpopulations is underway, and the status of the subpopulations should be reassessed once species status is clear.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Bennett N & da Silva JM. 2025. A conservation assessment of Georychus capensis. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The Cape Mole-rat is limited to the mesic regions of South Africa, mostly with annual rainfall levels above 500 mm (except in Nieuwoudtville, Citrusdal, Moorreesburg and Worcester). It has a discontinuous distribution across the south-western regions of South Africa. Its range extends northwards to Nieuwoudtville in the Northern Cape Province, and eastwards beyond Port Elizabeth to Bathurst. Fossil evidence suggests that it once had a much wider distribution (Hendey 1969, Klein 1974, Avery 1998), which contracted during the Quaternary (Klein 1974, Avery 1991). Populations in Mpumalanga and KwaZulu-Natal Provinces might therefore be geographical relicts (Avery 1991).
There are several additional isolated subpopulations, in southern KwaZulu-Natal along the border of Lesotho (specifically Nottingham Road and Impendle) and across central Mpumalanga (specifically Belfast, Ermelo and Wakkerstroom). The Tafelkop locality (Wakkerstroom) is a known subpopulation, but field surveys are required to locate new subpopulations in the Wakkerstroom area (N.C. Bennett, pers comm 2002; L. Cohen pers. comm. 2016). The presence of the subpopulation in Belfast was confirmed in 2015.
The species has not been recorded from Lesotho (Lynch 1994). Previous research suggests that this species once ranged along South Africa’s east coast (Avery 1991, Taylor 1998). Estimated extent of occurrence is 6,989 km² and 5,725 km² for the KZN and Mpumalanga subpopulations respectively.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 10m
Elevation Upper Limit (in metres above sea level): 1500-2000m
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
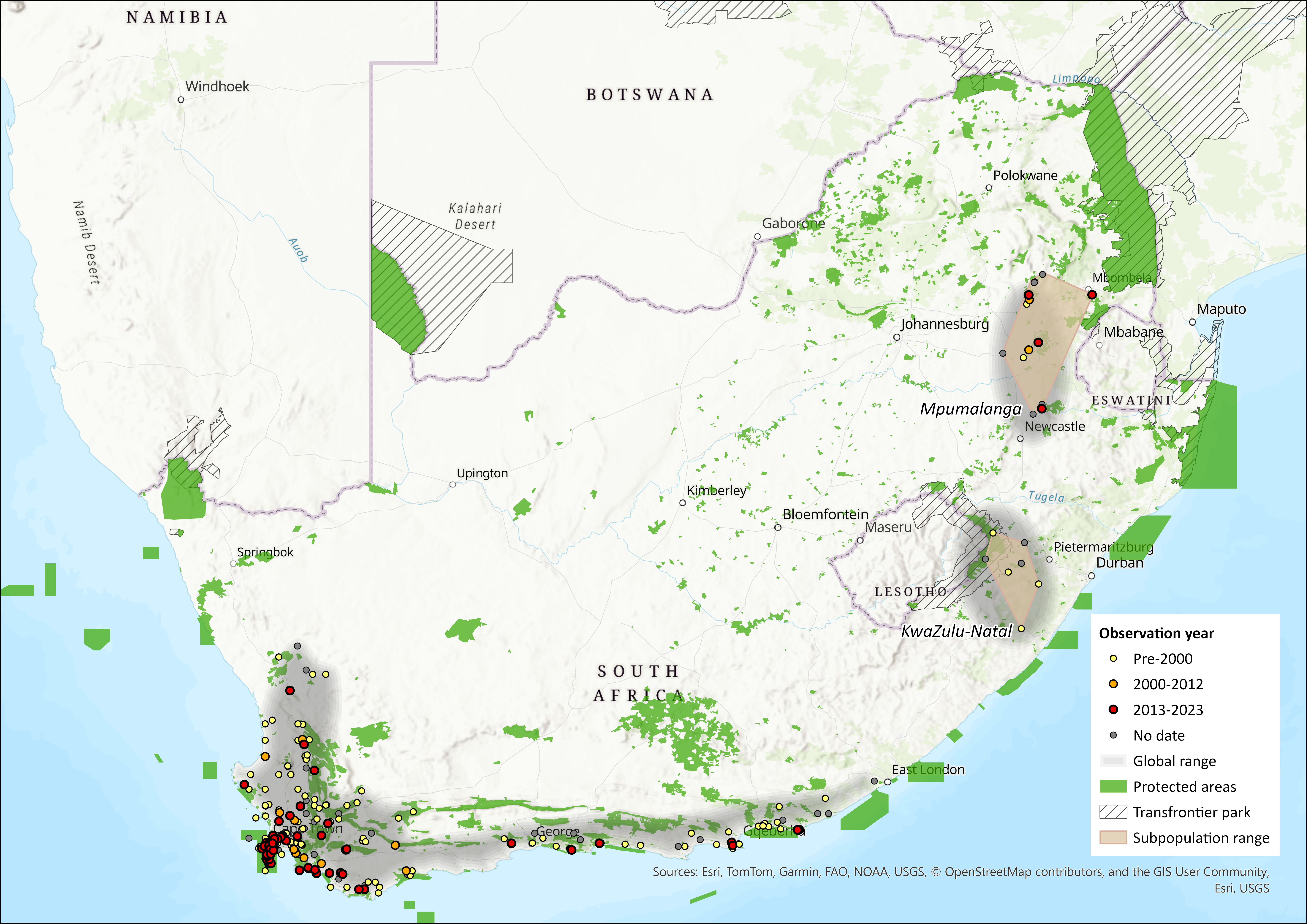
Figure 1. Distribution records for Cape Mole Rat (Georychus capensis) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| South Africa | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Climate change may threaten the fragmented subpopulations by reducing suitable habitat in the future. It could directly affect food resources and impact distribution in the areas bordering semi-arid areas (Hulme et al. 2001, Midgley et al. 2002).
Population information
The Cape Mole-rat is considered to be a common, localised species, with population densities occasionally exceeding 30 individuals / km² in the Cape Town region (N.C. Bennett unpubl. data). Naturally fragmented subpopulations have been identified in montane habitats of KwaZulu-Natal and Mpumalanga. Areas outside the known localities remain poorly sampled and thus it is possible that intermediate subpopulations exist which have not been discovered.
Current population trend: Stable (G. capensis); unknown (G. capensis – KZN and Mpumalanga subpopulations).
Population Information
Continuing decline in mature individuals? KwaZulu-Natal population is declining due to land development
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
All individuals in one subpopulation: No
Number of mature individuals in largest subpopulation: 100’s
Number of subpopulations: Many
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: Low
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: Low
Probability of extinction in the wild within 100 years: Medium to high due to climate change
Population genetics
The subpopulations from KwaZulu-Natal (Honeycutt et al. 1987, Nevo et al. 1987, J.H. Visser unpubl. data) and Mpumalanga (C.G. Faulkes and N.C. Bennett unpubl. data; J.H. Visser unpubl. data) are genetically distinct from each other, and ongoing molecular research is uncovering at least two lineages in the Western Cape. While population densities may be high in suitable areas, a detailed look into the abundances within each subpopulation have not been quantified. Consequently, an inference of the effective population sizes (Ne) of each and whether they exceed the accepted threshold of Ne 500 for genetically healthy and stable populations is not possible at this time.
Habitats and Ecology
The Cape Mole-rat prefers deep, sandy and clay soils, particularly coastal dunes and sandy alluvial deposits along rivers and montane areas of the Western Cape, Eastern Cape (Skinner and Chimimba 2005) and Mpumalanga (Visser et al. 2016) provinces. Generally, G. capensis is located in mesic areas receiving over 500 mm of rainfall per year. This species is commonly recorded in human-modified environments, such as golf courses, gardens and even at the main campus of the University of Cape Town. However, it is thought to be a habitat specialist that requires areas with vleis or in close proximity to rivers (Visser et al. 2016). Similarly, breeding pattern linked to seasonal rainfall, possibly resulting in distinct breeding seasons for different subpopulations (Visser et al. 2016). In the Cape, this species is found sympatrically with the Common Mole-rat (Cryptomys hottentotus). The subpopulation in Mpumalanga has been recorded in montane grassland areas consisting of red clay and black turf soils (Bronner 1990), and loose or sandy soils around pans (Roberts 1951). The subpopulation in KwaZulu-Natal Province occurs within mesic grasslands on sandy or clay soils, particularly in the midlands (for example, Impendle and Nottingham Road). Overall, the most commonly preferred soil types are derived from sandstone, limestone, shale and quartzite which comprise the sandy loam, clay and alluvium soils that are inhabited (Visser et al. 2016). Predominantly solitary, the Cape Mole-rat produces two litters of between four and ten young each year (Bennett and Jarvis 1988), with a generation length of four years (N.C. Bennett unpubl. data). They are solitary and burrow about 200 mm below the surface (Skinner and Chimimba 2005), and may store food, such as geophytes, in underground chambers (du Toit et al. 1985). They mainly eat plant storage organs. The seismic signal consists of a series of long bursts of very rapid drums (Bennett and Jarvis 1988).
As excellent soil engineers, the burrowing activities of subterranean mole-rats can lead to modification of soil properties, thus enhancing the species richness of vegetation communities (Hagenah and Bennett 2013). Specifically, in the Cape, soils of Cape Mole-rat mounds were found to have higher nitrogen and magnesium levels, when compared to soils unaffected by Mole-rats (Hagenah and Bennett 2013). An increase in species richness, as a result subterranean Mole-rats in the Cape Floristic Region may be particularly important considering the high conservation value of this area. Additionally, they create refuges for other species to use when escaping fire, and limit surface runoff of water by lowering the compactness of soil thus increasing drainage and moisture-holding capacity (Hagenah and Bennett 2013).
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 1.4. Forest -> Forest – Temperate | – | Suitable | – |
| 3.8. Shrubland -> Shrubland – Mediterranean-type Shrubby Vegetation | – | Suitable | – |
| 14.1. Artificial/Terrestrial -> Artificial/Terrestrial – Arable Land | – | Suitable | – |
| 14.4. Artificial/Terrestrial -> Artificial/Terrestrial – Rural Gardens | – | Suitable | – |
| 14.5. Artificial/Terrestrial -> Artificial/Terrestrial – Urban Areas | – | Suitable | – |
Life History
| Generation Length | Justification | Data Quality |
| 4 | – | good |
Age at Maturity: Female or unspecified: 1 year
Age at Maturity: Male: 1 year
Size at Maturity (in cms): Female: 21cm 181g
Size at Maturity (in cms): Male: 20cm 180g
Longevity: 6-8 years
Average Reproductive Age: 20cm, 1 year
Maximum Size (in cms): 23cm
Size at Birth (in cms): 2cm, 5-12g
Gestation Time: 44-48 days
Reproductive Periodicity: Seasonal two litters per annum
Average Annual Fecundity or Litter Size: 6 (4-12)
Natural Mortality: Mole snake, cats, dogs
Does the species lay eggs? No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage? No
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: Dispersal above ground
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species is not traded or utilised in any way, aside from limited numbers of individuals collected for laboratory research, but this has no impact on the population. Georychus capensis is not used to supplement protein intake by humans.
Local Livelihood: No
National Commercial Value: No
International Commercial Value: No
End Use: None
Is there harvest from captive/cultivated sources of this species? Research at universities
Harvest Trend Comments: n/a
Threats
No major threats to this species have been identified in the Cape. The threats associated with loss in habitat quantity and quality due to the development of timber plantations and afforestation in the midlands and montane grasslands of KwaZulu-Natal and Mpumalanga have been stabilized (no further plantations planned) and are no longer regarded as major dangers to this species. However, afforestation makes the soils on which this species depends less suitable (Armstrong et al. 1998) and has thus effectively reduced area of occupancy. The isolated subpopulations in Mpumalanga may face increasing threats of habitat loss and degradation as a result of mining, agriculture, and human settlements where 77% of the grassland patches are <10 km² in size (Neke and du Plessis 2004). The KwaZulu-Natal subpopulation is very rarely seen in gardens, which suggests it will not adapt to human-modified landscapes. This species faces an additional threat of predation by dogs in urban and some rural areas as well as persecution by farmers and homeowners who view this species as a pest, particularly within small scale vegetable gardens. Overall, ongoing habitat loss and fragmentation from agricultural, human settlement and industrial expansion, combined alien vegetation infestation and incorrect fire regimes may be causing a decline into the two subpopulations.
Overall, there was a 20.4% loss of natural habitat from 1994 to 2011, with an average loss of 1.2% per annum, due primarily to agriculture (5.2% increase; 4,962 km²), but also plantations, built environments and settlements, mines and dams (Jewitt et al. 2015). There has been no analysis of rate of loss within Mpumalanga Province but the Mpumalanga Tourism and Parks Agency (MTPA) mapped all development applications received at a cadastral scale over a 14-year period (2000-2014), which showed that greatest pressure for land-use change has come from prospecting applications (54% of the land surface area) and mining (25% of land surface area), with the balance of applications related to built infrastructure (9%), residential development (4%) and cultivation (0.7%) (Lötter et al. 2014). Future developments at the above rates or even higher are likely to cause further detriment towards natural ecosystems and processes that this species is associated with.
However, it should be noted that anthropogenic disturbances do not necessarily cause population declines or a loss of habitat as grazing and the planting of lawns creates suitable habitat and they are found in such landscapes across the distribution. Apparent range contractions in certain (broad) areas of KwaZulu-Natal and Mpumalanga may constitute natural fragmentation as a result of the life-history of the animal being sessile and subterranean, along with its specialised habitat type, and may have become occurred historically before human influence.
Conservation
The Cape Mole-rat occurs in several key protected areas within the assessment region, such as Table Mountain National Park, Cederberg Wilderness Area and Langebaan Nature Reserve in the Western Cape. Additionally, the Wakkerstroom subpopulation in Mpumalanga occurs on the farm Tafelkop 126 HT, which has been recently declared a nature reserve and is known as the Tafelkop Nature Reserve (L. Cohen pers. comm. 2016). The Mpumalanga Biodiversity Sector Plan (MBSP) has been developed by the Mpumalanga Tourism and Parks Agency (MTPA) and is based on a systematic conservation plan, which considers the distribution of all species and their habitats. According to the MBSP, large parts of the farm where the known Belfast subpopulation occurs are situated in an irreplaceable and optimal Critical Biodiversity Area that has high value of conservation status. The KwaZulu-Natal subpopulation, however, only occurs in the Drakensberg foothills and not the protected areas; and should thus be prioritised for protected area expansion.
The following interventions are encouraged:
- Work in local communities to prevent poisoning and persecution.
- Educate and train farmers to mitigate poor grazing and burning regimes. For example, in Mpumalanga Province, structured veld management training programmes are provided for formally proclaimed areas which form part of protected area expansion/stewardship initiatives.
- Expand protected areas to preserve the remaining habitat for the KZN and Mpumalanga subpopulations.
Finally, further studies into the taxonomic status and distributions of the KwaZulu-Natal and Mpumalanga subpopulations are needed. If the subpopulations in Mpumalanga and KwaZulu-Natal are conclusively found to be of different taxonomic status (as is suspected) then they should be reassessed. These two subpopulations are characterised by a small geographic area low densities of occurrence (except in Wakkerstroom).
Recommendations for land managers and practitioners:
- Systematic surveys and ongoing monitoring to document subpopulation localities and densities.
- Reduce overgrazing and implement correct fire regime.
Research priorities:
- Further studies into the taxonomic status of the KZN and Mpumalanga subpopulations are needed.
- Studies assessing the population size, trend and distribution, particularly of KZN and Mpumalanga subpopulations.
Encouraged citizen actions:
- Report sightings to conservation officials and researchers and on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Deposit any dead specimens at your local conservation office for identification.
Bibliography
Armstrong AJ, Benn G, Bowland AE, Goodman PS, Johnson DN, Maddock AH, Scott-Shaw CR. 1998. Plantation forestry in South Africa and its impact on biodiversity. The Southern African Forestry Journal 182: 59-65.
Aver, D.M. 1991. Late Quaternary incidence of some micromammalian species in Natal. Durban Museum Novitates 16: 1–11.
Avery, D.M. 1998. An assessment of the lower pleistocene micromammalian fauna from Swartkrans members 1–3, Gauteng, South Africa. Geobios 31: 393–414.
Bennett, N.C. and Jarvis, J.U.M. 1988. The reproductive biology of the Cape mole-rat, Georychus capensis (Rodentia, Bathyergidae). Journal of Zoology 214: 95–106.
Bronner, G.N. 1990. New distribution records for four mammal species, with notes on their taxonomy and ecology. Koedoe 33: 1-7.
Bennett, N.C., Maree, S. & Faulkes, C.G. (2006). The Cape blesmol, Georychus capensis. Mammalian Species 799:1-4.
Hagenah, N. and Bennett, N.C. 2013. Mole rats act as ecosystem engineers within a biodiversity hotspot, the Cape Fynbos. Journal of Zoology 289: 19-26.
Hendey, Q.B. 1969. Quaternary vertebrate fossil sites in the south-western Cape Province. The South African Archaeological Bulletin 24: 96–105.
Honeycutt, R.L., Edwards, S.V., Nelson, K. and Nevo, E. 1987. Mitochondrial DNA variation and the phylogeny of African mole-rats (Rodentia: Bathyergidae). Systematic Zoology 36: 280–292.
Hulme, M., Doherty, R., Ngara, T., New, M. and Lister, D. 2001. African climate change: 1900-2100. Climate Research 17: 145-168.
IUCN. 2017. The IUCN Red List of Threatened Species. Version 2017-2. Available at: www.iucnredlist.org. (Accessed: 14 September 2017).
Jewitt, D., Goodman, P.S., Erasmus, B.F.N., O’Connor, T.G. and Witkowski, E.T.F. 2015. Systematic land-cover change in KwaZulu-Natal, South Africa: Implications for biodiversity. South African Journal of Science 111: 1-9.
Klein, R.G. 1974. A provisional statement on terminal Pleistocene mammalian extinctions in the Cape biotic zone (Southern Cape Province, South Africa). South African Archaeological Society, Goodwin Series 2: 39–45.
Lynch, C.D. 1994. The mammals of Lesotho. Navorsinge van die Nasionale Museum Bloemfontein 10(4): 177-241.
Lötter, M.C., Cadman, M.J. and Lechmere-Oertel, R.G. 2014. Mpumalanga Biodiversity Sector Plan Handbook. MTPA. Nelspruit.
Midgley, G.F., Hannah, L., Millar, D., Rutherford, M.C. and Powrie, L.W. 2002. Assessing the vulnerability of species richness to anthropogenic climate change in a biodiversity hotspot. Global Ecology and Biogeography 11: 445–451.
Narins, P.M., Reichman, O.J., Jarvis, J.U.M., Lewis, E.R. 1992. Seismic signal transmission between burrows of the Cape mole-rat, Georychus capensis. Journal of Comparative Physiology A 170: 13–21.
Neke, K.S. and du Plessis, M.A. 2004. The threat of transformation: quantifying the vulnerability of grasslands in South Africa. Conservation Biology 18: 466–477.
Nevo, E., Ben-Shlomo, R., Beiles, A., Jarvis, J.U.M. and Hickman, G.C. 1987. Allozyme differentiation and systematics of the endemic subterranean mole rats of South Africa. Biochemical Systematics and Ecology 15: 489-502.
Roberts, A. 1951. The Mammals of South Africa. Central New Agency, Johannesburg, South Africa.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Taylor, P.J. 1998. The Smaller Mammals of KwaZulu-Natal. University of Natal Press, Pietermaritzburg, South Africa.
Visser, J.H., Bennett, N.C. and van Vuuren, B.J. 2016. Distributional range, ecology and mating system of the Cape mole-rat, Georychus capensis family Bathyergidae. Canadian Journal of Zoology.
du Toit, J.T., Jarvis, J.U.M. and Louw, G.N. 1985. Nutrition and burrowing energetics of the Cape mole-rat Georychus capensis. Oecologia 66: 81–87.