

Bushveld Gerbil
Gerbilliscus leucogaster

2025 Red list status
Least Concern
Regional Population Trend
Stable
Change compared
to 2016
No Change
Overview
Gerbilliscus leucogaster – (Peters, 1852)
ANIMALIA – CHORDATA – MAMMALIA – RODENTIA – MURIDAE – Gerbilliscus – leucogaster
Common Names: Bushveld Gerbil (English), Bosveldse Nagmuis (Afrikaans)
Synonyms: Tatera leucogaster (Peters, 1852); see Dempster (2013) for full list.
Taxonomic Note:
Previously classified as Tatera leucogaster with at least 17 described subspecies in southern Africa (Skinner & Chimimba 2005), this species was later recognised as Gerbilliscus leucogaster by Musser and Carleton (2005). The Bushveld Gerbil can be distinguished in its range from other Gerbilliscus species by its long tail and the black dorsal line, which runs along the tail length (Monadjem et al. 2015).
Red List Status: LC – Least Concern
Assessment Information
Assessors: Smith, C.1 & da Silva, J.M.2
Reviewers: Taylor, P.J.3
Contributor: Patel, T.1
Institutions:1Endangered Wildlife Trust,2South African National Biodiversity Institute,3University of the Free State
Previous Assessors and Reviewers: du Plessis, J., Swanepoel, L.H., McDonough, M. & Shoeman, C.
Previous Contributors: Child, M.F., Avenant, N., Avery, M., Baxter, R., MacFadyen, D., Monadjem, A., Palmer, G., Taylor, P. & Wilson, B.
Assessment Rationale
Listed as Least Concern in view of its wide distribution within the assessment region, abundant population, ability to live in a wide range of habitats, including agricultural landscapes, and because there are no major threats that could cause population decline. It may become an agricultural pest on grain-growing lands during some periods where populations can rapidly increase under optimal food conditions, which could result in local persecutions. Selective chemical control or biocontrol of population explosions through the use of Barn Owls (Tyto alba) are potentially effective and target-specific methods that are currently available and should be encouraged as techniques for holistic management .
Regional population effects: Rescue effect is possible through dispersal from Namibia, Botswana, Zimbabwe and Mozambique. Immigration is unlikely to decrease.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended Citation: Smith C & da Silva JM. 2025. A conservation assessment of Gerbilliscus leucogaster. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
Endemic to Africa, the global distribution for this species ranges from Tanzania in the north to South Africa and from eastern Namibia and Angola to western and northern Mozambique. It is widely distributed in the Zambezian Woodland biotic zone and in parts of the South-West Arid and Highveld biotic zones (Dempster 2013). It occurs widely from the equator to about 30°S, including the lowveld of Swaziland and northeastern KwaZulu-Natal (Dempster 2013). It occurs in the northern and central parts of Namibia, throughout Botswana and Zimbabwe and in Mozambique south of the Zambezi River (Skinner & Chimimba 2005).
In the assessment area, it occurs in South Africa throughout the North West and Limpopo provinces (Skinner & Chimimba 2005; Power 2014), in most of Mpumalanga (except the south-central parts; Skinner & Chimimba 2005) and in the southern and western parts of the Free State (Lynch 1983; Skinner & Chimimba 2005). In the Northern Cape, most records are restricted to a strip to the north of the Orange River (Skinner & Chimimba 2005) although Avery and Avery (2011) have listed records spanning further westwards. It is generally restricted to the northeastern parts of KwaZulu-Natal (Skinner & Chimimba 2005), with Avery et al. (2002) reporting a slight range expansion for these species to the south, but these records are still to be verified (see Monadjem et al. 2015). It occurs in the northern half of Eswatini (Monadjem 1997a; Skinner & Chimimba 2005) and is currently considered to be absent from Lesotho (Lynch 1994; Ambrose 2006). In the Free State Province, this species has been recorded from Tussen-die-Riviere Nature Reserve (Ferreira & Avenant 2003).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
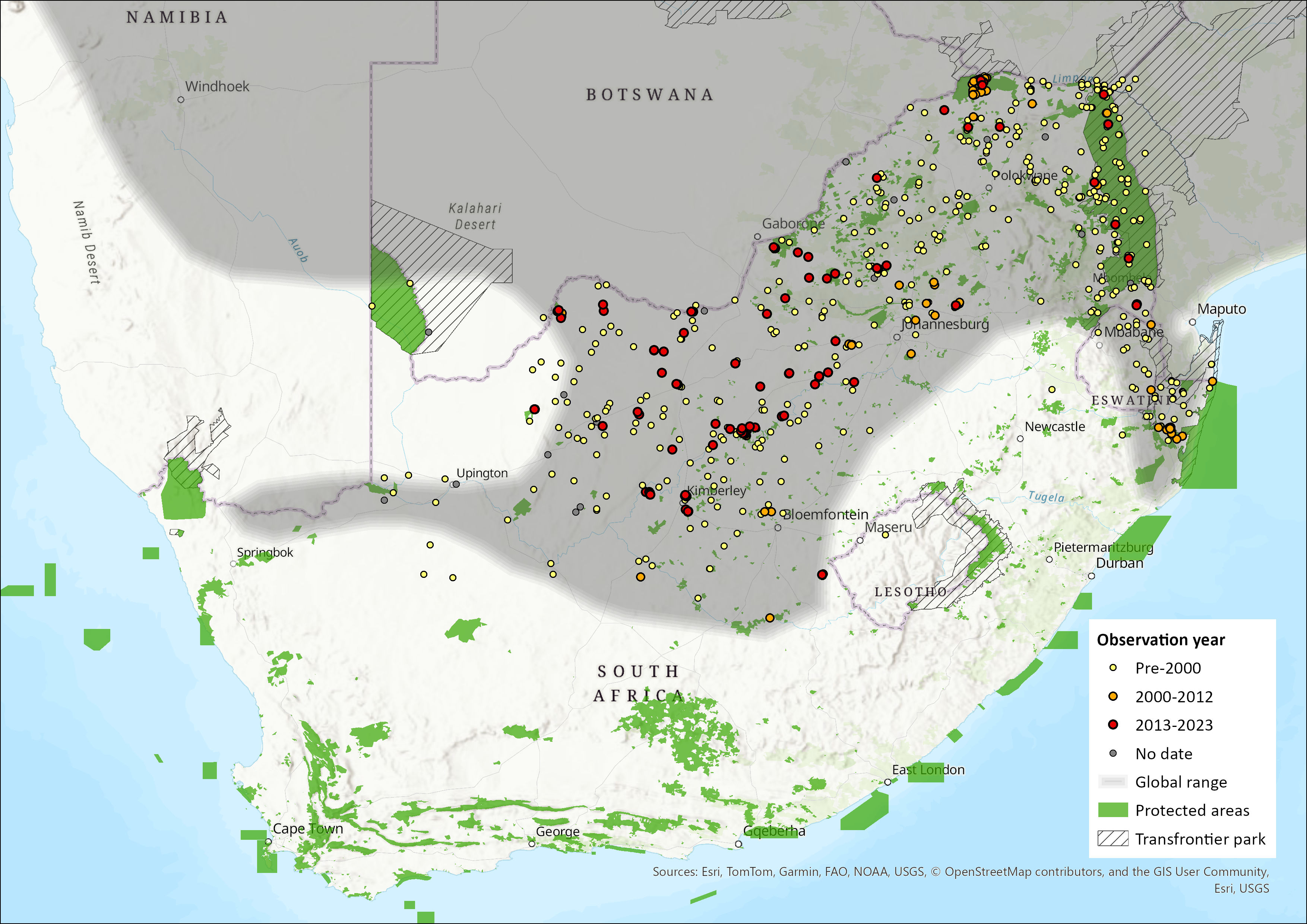
Figure 1. Distribution records for Bushveld Gerbil (Gerbilliscus leucogaster) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Lesotho | Possibly Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
No recent research or literature has focused on how climate change might affect this species in the future.
Population
This species is relatively common, exhibiting expected cyclic fluctuations of population abundance (Avenant 2011; Dempster 2013). It is often the most common small mammal species in an area (Linzey & Kesner 1997; Power 2014). Its population numbers and frequency of occurrence may vary with habitat or environmental conditions. It was the most common species in habitats that were frequently burned, compared to unburned habitat, in the Kruger National Park (Korn 1981) and Nysvley (Korn 1987). In the Okavango Delta, Plavsic (2014) found that numbers in an unburned site were marginally higher compared to numbers in a burned site for six months post-burning, thereafter the pattern was reversed. In small-holder agricultural systems of the Vhembe District, Limpopo Province, it reached higher abundances in the wetter areas (Vyeboom village, minimum number alive – 30 animals / ha), than the dryer areas (Ka-Ndengeza villages, minimum number alive – 11 animals / ha; L.H. Swanepoel, L.N. Nembudani & P.J. Taylor unpubl. data). In commercial agro-ecosystems of the Free State Province, populations can reach high densities (minimum number alive – up to 150 animals / ha; L.H. Swanepoel & M. Keith, unpubl. data), while in natural grassland (Sandveld Nature Reserve) it can be the dominant species (minimum number alive – 25 animals / ha; L.H. Swanepoel & M. Keith, unpubl. data). On the Highveld grasslands it often surprisingly outnumbers the Highveld Gerbil (G. brantsii) (Power 2014). At Nylsvley, abundance varied from 6% (in old fields; least common of the five species sampled) to 61% (in burnt Acacia woodland; most common of the five species sampled) (Korn 1987).
Population Information
Current population trend: Stable
Continuing decline in mature individuals: Unknown
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: Unknown
Severely fragmented: No
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
A phylogeographic study incorporating both mitochondrial and nuclear (AFLP) markers uncovered extensive genetic structure within this species across its range (McDonough et al. 2015). Three mitochondrial clades and two nuclear clades were found within the assessment region; however, it must be noted that samples from the third clade were not incorporated in the nuclear analysis. Consequently, it is possible three clades are present, which could correspond to evolutionary significant units (ESUs), or at the very least subpopulations.
The three clades correspond to:
- northern South Africa (mtDNA clade A, nuclear: central clade)
- Northern Cape/North-West provinces (mtDNA clade D, nuclear: southern clade)
- northern KwaZulu-Natal (mtDNA clade B, nuclear: unknown)
- To verify these clades, a more fine-scale population genomic study incorporating more comprehensive sampling across the assessment region should be undertaken. This study would also be influential in understanding the processes and features responsible for the strong molecular structure despite seemingly continuous distribution (i.e., vicariance) for this species.
Given the species is thought to be abundant across its range, it is expected that great than 5 000 mature individuals make up each subpopulation; thereby exceeding the Ne 500 threshold.
Based on the available information, the two genetic indicators adopted in the Convention of Biological Diversity’s Global Biodiversity Framework can be quantified:
- Proportion of populations maintained (PM) indicator: 3 extant populations remain, no known extinctions; 3/3 = 1.0.
- Proportion of populations with an effective population size [Ne] greater than 500: all assumed to exceed 500; 3/3 = 1.0
Habitats and ecology
It is generally associated with a wide variety of habitats, including bushveld and grasslands (Skinner & Chimimba 2005) and may even occur in highly transformed habitats (Power 2014), typically occurring in areas with mean annual rainfall above 250 mm (Dempster 2013). In the North West Province, it is found in over half of the province’s vegetation types, and commonly colonises the bare understory of exotic stands of Prosopis glandulosa (Power 2014). Although it is predominantly associated with light sandy soils or sandy alluvium, it may also occur on hard ground where it uses holes in termitaria and under tree roots (Skinner & Chimimba 2005; Dempster 2013). It is absent in areas of heavy red clay soils or soft sand (Dempster 2013). In agro-ecosystems dominated by sandveld, it seems to co-occur with G. brantsii, where it seems to outcompete them (L.H. Swanepoel and M. Keith unpubl. data). The frequency and timing of burning can have a significant effect on its abundance. Generally, the highest proportions of communities occur in habitats that have been burnt one or more times during the previous three years, compared to unburnt sites (Korn 1981). In KwaZulu-Natal, this species has been found in Acacia nilotica/Hyphaene coriacae pan systems, H. coriacae Palmveld, Sand forest, Combretum molle woodland and riverine woodland (Rautenbach et al. 2014; Delcros et al. 2015).
It has an omnivorous diet, which mainly consists of plant material and insects (Perrin & Swanepoel 1987; Monadjem 1997b). In some parts of South Africa, it may potentially cause significant damage in cropland when it forages on germinating seeds and newly emergent seedlings (Verdoorn 2010; von Maltitz et al. 2014). It is often the most abundant small mammal species in agricultural areas (L.H. Swanepoel & M. Keith unpubl. data), where it can reach excessively high numbers during population explosions (de Graaff 1981).
They are nocturnal and lives in groups or in pairs (Pinto 2023). This species excavates small burrows, which are interconnected underground, and cleans its burrows every evening. As such, burrows in use can be recognised by fresh soil outside the burrow (Skinner & Chimimba 2005; Dempster 2013). This species prefers deep and light sandy soils (Starik 2020). Breeding is seasonal and associated with regular summer rainfall. The duration of the breeding season is variable, which is indicative of a temporally dynamic reproductive strategy. Litter size is large but variable and associated with a xeric environment (Perrin & Swanepoel 1987).
Ecosystem and cultural services: It has been recorded as a reservoir of the plague bacillus Yersinia pestis in South Africa, and is also susceptible to infection of African horse sickness and Listeria monocytogenes under laboratory conditions (Dempster 2013). With the exception of periods during which this species exhibits low population numbers, it is also a valuable food source for small carnivores and raptors, especially Barn Owls. G. leucogaster can be used as an ecological indicator of ecosystem integrity as well as the condition of savanna landscapes (Starik 2020).
IUCN Habitats Classification Scheme
|
Habitat |
Season |
Suitability |
Major Importance? |
|
2.1. Savanna -> Savanna – Dry |
– |
Suitable |
– |
|
3.4. Shrubland -> Shrubland – Temperate |
– |
Suitable |
– |
|
4.5. Grassland -> Grassland – Subtropical/Tropical Dry |
– |
Suitable |
– |
|
8.2. Desert -> Desert – Temperate |
– |
Suitable |
– |
Life History
Generation Length: (Not specified)
Age at maturity: female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species is not known to be traded or utilised in any form.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
There are no major threats to this species. During population explosions it may become an agricultural pest in some areas, which could result in persecution (Verdoorn 2010; von Maltitz et al. 2014). Pest control depends on the farming set-up, but mostly takes the form of rodenticides, chemical seed treatment or habitat modification. These control actions can have negative effects on other non-pest rodent species (Makundi & Massawe 2011), as well as the secondary predators feeding on rodents (Taylor et al. 2012). This species may be a reservoir for the bubonic plague (Dempster 2013), however, it is unlikely that it is persecuted for this, as it does not usually enter human establishments. This species shows a sensitivity to land use-related soil degradation (Starik 2020).
Current habitat trend: Stable
Conservation
This species is present in many protected areas within the assessment region, for example, Mkhuze Game Reserve, Phinda Private Game Reserve, Kruger National Park, and healthy populations are known to exist in the Sandveld Nature Reserve, Free State (L.H. Swanepoel & M. Keith, unpubl. data). While no specific interventions seem to be necessary for its conservation, careful attention should be paid to the secondary impact of pest management. It is suggested that management of pest Highveld Gerbils be done using ecologically-based rodent management principles (Taylor et al. 2012). These include responsible use of chemicals, rodenticides, natural predation (biocontrols), habitat modification and intensive communal trapping (Makundi & Massawe 2011). In a study conducted on G. afra in the Western Cape, Barn Owl control was found to be twice as effective for controlling the gerbil population, compared to the use of poison (Potter 2004). Barn Owls respond to prey explosions by increasing reproduction (Potter 2004; Makundi & Massawe 2011). This management intervention also saves farmers the cost of buying poison and prevents the mortality of non-target species, including domestic stock.
Recommendations for land managers and practitioners:
- Although the likelihood of success should still be established for G. leucogaster, Barn Owls could potentially be used to control population explosions of this species in cropland. This should be encouraged as part of a holistic management intervention (Verdoorn 2010). To increase the potential utility of Barn Owls to manage G. leucogaster, perches should be erected in crop fields and nesting boxes in appropriate sites close to the fields.
Research priorities:
- The practicalities and effectiveness of the use of Barn Owls as a population control method for this species.
- Taxonomic resolution of the Gerbilliscus genus.
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Install Barn Owl nest boxes and perches in crop fields to encourage biocontrol during population explosions.
Bibliography
Ambrose D. 2006. Lesotho annotated bibliography. Section 168: mammals including annotated species lists. House 9 Publications, National University of Lesotho, Roma.
Avenant N. 2011. The potential utility of rodents and other small mammals as indicators of ecosystem ’integrity’ of South African grasslands. Wildlife Research 38: 626–639.
Avery DM, Avery G. 2011. Micromammals in the Northern Cape Province of South Africa, past and present. African Natural History 7: 9–39.
Avery, D.M., Avery, G. and Roberts, A. 2002. A contribution from barn owl pellets to known micromammalian distributions in KwaZulu-Natal, South Africa. African Zoology 37: 131-140.
De Graaff, G. 1981. The Rodents of Southern Africa. Butterworths, Durban, South Africa.
Delcros G, Taylor PJ, Schoeman MC. 2015. Ecological correlates of small mammal assemblage structure at different spatial scales in the savannah biome of South Africa. Mammalia 79: 1–14.
Dempster ER. 2013. Gerbilliscus leucogaster Bushveld gerbil. Pages 279–281 in Happold DCD, editor. Mammals of Africa: Volume III: Rodents, Hares and Rabbits. Bloomsbury Publishing, London, UK.
Ferreira, S.M. and Avenant, N.L. 2003. Modelling the effects of trap-spacing on small mammal community descriptors in grasslands at Tussen-die-Riviere Nature Reserve, Free State Province, South Africa. Navorsinge van die Nasionale Museum, Bloemfontein 19: 21-30.
Korn H. 1987. Densities and biomasses of non-fossorial southern African savanna rodents during the dry season. Oecologia 72: 410 –413.
Korn NG. 1981. The influence of fire on populations of small mammals of the Kruger National Park. Koedoe 24: 125–157.
Linzey AV, Kesner MH. 1997. Small mammals of a woodland savannah ecosystem in Zimbabwe. I. Density and habitat occupancy patterns. Journal of Zoology (London) 243: 137–152.
Lynch CD. 1994. The mammals of Lesotho. Navorsinge van die Nasionale Museum, Bloemfontein 10: 177–241.
Lynch, C.D. 1983. The mammals of the Orange Free State, South Africa. Navorsinge van die Nasionale Museum Bloemfontein 18: 1-218.
Makundi RH, Massawe AW. 2011. Ecologically based rodent management in Africa: potential and challenges. Wildlife Research 38: 588–595.
McDonough, M., Šumbera, R., Mazoch, V., Ferguson, A., Phillips, C & Bryja, J. 2015. Multilocus phylogeography of a widespread savanna-woodland adapted rodent reveals the influence of Pleistocene geomorphology and climate change in Africa’s Zambezi region. Molecular Ecology. 24.10.1111/mec.13374
Monadjem A. 1997. Habitat preferences and biomasses of small mammals in Swaziland. African Journal of Ecology 35: 64-72.
Monadjem A. 1997. Stomach contents of 19 species of small mammals from Swaziland. South African Journal of Zoology 32: 23–26.
Monadjem, A., Taylor, P.J., Denys, C. and Cotterill, F.P.D. 2015. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis. . De Gruyter, Berlin, Germany.
Musser, G.G. and Carleton, M.D. 2005. Superfamily Muroidea. In: D.E. Wilson and D.A. Reeder (eds), Mammal Species of the World: a geographic and taxonomic reference, pp. 894-1531. The John Hopkins University Press, Baltimore, USA.
Perrin MR, Swanepoel P. 1987. Breeding biology of the bushveld gerbil Tatera leucogaster in relation to diet, rainfall and life history theory. South African Journal of Zoology 22: 218–227.
Pinto, C.J., 2023. Assessing habitat selection of grassland rodents in the Cradle of Humankind.
Plavsic MJ. 2014. Proximate and ultimate drivers of smallmammal recolonization after fire: microhabitat conditions, rainfall and species traits. Animal Conservation 17: 573–582.
Potter L. 2004. Raptors for rodent control: Is the Barn Owl a viable control agent for pest rodents on South African farmlands? University of Cape Town.
Power RJ. 2014. The Distribution and Status of Mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng, South Africa.
Rautenbach A, Dickerson T, Schoeman MC. 2014. Diversity of rodent and shrew assemblages in different vegetation types of the savannah biome in South Africa: no evidence for nested subsets or competition. African Journal of Ecology 52: 30-40.
Skinner J.D. and Chimimba C.T. 2005. The Mammals of the Southern African Subregion. Cambridge University Press, Cambridge, UK.
Starik, N., Mbango, O.K., Bengsch, S., Göttert, T. and Zeller, U., 2020. Landscape transformation influences responses of terrestrial small mammals to land use intensity in north-central Namibia. Diversity, 12(12), p.488.
Taylor PJ, et al. 2012. Experimental treatment-control studies of ecologically based rodent management in Africa: balancing conservation and pest management. Wildlife Research 39: 51–61.
Verdoorn GH. 2010. Environmentally compatible rodent management for agriculture. Griffon Poison Information Centre.
von Maltitz E, Kirsten F, Malebana P. 2014. Update on gerbil management in maize. SA Grain 16: 106–107.