

Banded Mongoose
Mungos mungo

2025 Red list status
Least Concern
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Mungos mungo – (Gmelin, 1788)
ANIMALIA – CHORDATA – MAMMALIA – CARNIVORA – HERPESTIDAE – Mungos – mungo
Common Names: Banded Mongoose, Zebra Mongoose (English), Gebande Muishond, Barasinga (Afrikaans), Usikibhoror (Ndebele), Moswe, Moswê (Sepedi), Letodi (Sesotho), Letôtôtô, Letara, Lejara (Setswana), Lichacha (Swati), Nkala (Tsonga), Tshihoho, Tzwikitowe (Venda), Ubuhala, Ubuhaye (Zulu)
Synonyms: Viverra mungo Gmelin, 1788
Taxonomic Note:
Although there is historic recognition of the southern African subspecies, Mungos mungo taenionotus (Kingdon 1997), from KwaZulu-Natal and Mpumalanga, and M. m. grisonax from the North West, Limpopo and Gauteng provinces (Skinner & Chimimba 2005), these subspecies are no longer recognised. Skinner and Chimimba (2005) describe the variation in pelage colour between these previously recognised subspecies, with M. m. grisonax lighter in colour than M. m. taenianotus. Recent molecular studies have shown that the Banded Mongoose is in the subfamily Mungotinae, along with several other social African mongooses (Veron et al. 2022).
Red List Status: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Power, R.J.1 & da Silva, J.M.2
Reviewer: Do Linh San, E.3
Contributor: Nicholson, S.K.4
Institutions: 1North West Provincial Government, 2South African National Biodiversity Institute, 3Sol Plaatjie University, 4Endangered Wildlife Trust
Previous Assessors and Reviewers: Gilchrist, J.S., Stuart, C., Stuart, M. & Do Linh San, E.,
Previous Contributor: Hoffmann, M.
Assessment Rationale
The Banded Mongoose is listed as Least Concern as, although its distribution is restricted to the northeast of the assessment region, it is generally common in suitable habitat and is present in several protected areas. There are no major threats that could cause range-wide population decline. Accidental persecution through poisoning, controlled burning, and infectious disease may lead to local declines, whilst wildlife ranching might have a positive effect by conserving more suitable habitat and connecting subpopulations.
Regional population effects: Dispersal across regional borders is suspected as the range extends widely into Mozambique and is continuous into southeastern Botswana and southern Zimbabwe, and the species is not constrained by fences (Gilchrist et al. 2016).
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Power RJ & da Silva JM. 2025. A conservation assessment of Mungos mungo. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
This species is distributed widely in sub-Saharan Africa from Senegal and Gambia to Ethiopia, Eritrea and Somalia, and south to about 31° in South Africa (Skinner and Chimimba 2005; Gilchrist et al. 2016). It has been recorded to 1,600 m above sea level in Ethiopia (Yalden et al. 1996). Although fairly widespread in southern Africa, M. mungo appears to be rare in West Africa. Its relative scarcity in West Africa may be due to niche overlap with its congener, the Gambian Mongoose (M. gambianus), endemic to West Africa and reported to occupy similar habitat and have a similar diet (Cant & Gilchrist 2013; van Rompaey & Sillero-Zubiri 2013).
Within the assessment region, Banded Mongooses occur in bushveld in Limpopo Province, Mpumulanga, Gauteng, North West Province, and the KwaZulu-Natal coastal plain (Gilchrist et al. 2016). They are also present as an apparently isolated population in the Kgalagadi Transfrontier Park, centred on the Nossob River, Northern Cape Province (C. Stuart & M. Stuart pers. obs. 2000, In: Gilchrist et al. 2016), though this may likely be a consequence of sporadic occurrences, and less so a resident population. It is possibly linked to either the northern population, but information is lacking.
In the North West, the species occurs in the northern Bushveld (Power et al. 2019). In 2021, the species is reported to still be seen in the western end of the range in the assessment region (Figure 1). It still occurs across the northern provinces from Gauteng, Limpopo and Mpumalanga, in particular the Lowveld Reserves (Smith et al. 2023). In KwaZulu-Natal, the species occurs in the Zululand reserves (Da Rosa 2019; Pretorius 2019), and occurs along the coast toward Durban (Figure 1). There appears to be no recent records in north-west KwaZulu-Natal up the Pongola valley (Figure 1).
Although uncommon, the species also occurs in Eswatini (Monadjem 1998) and seems still present (Figure 1).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): (Not specified)
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
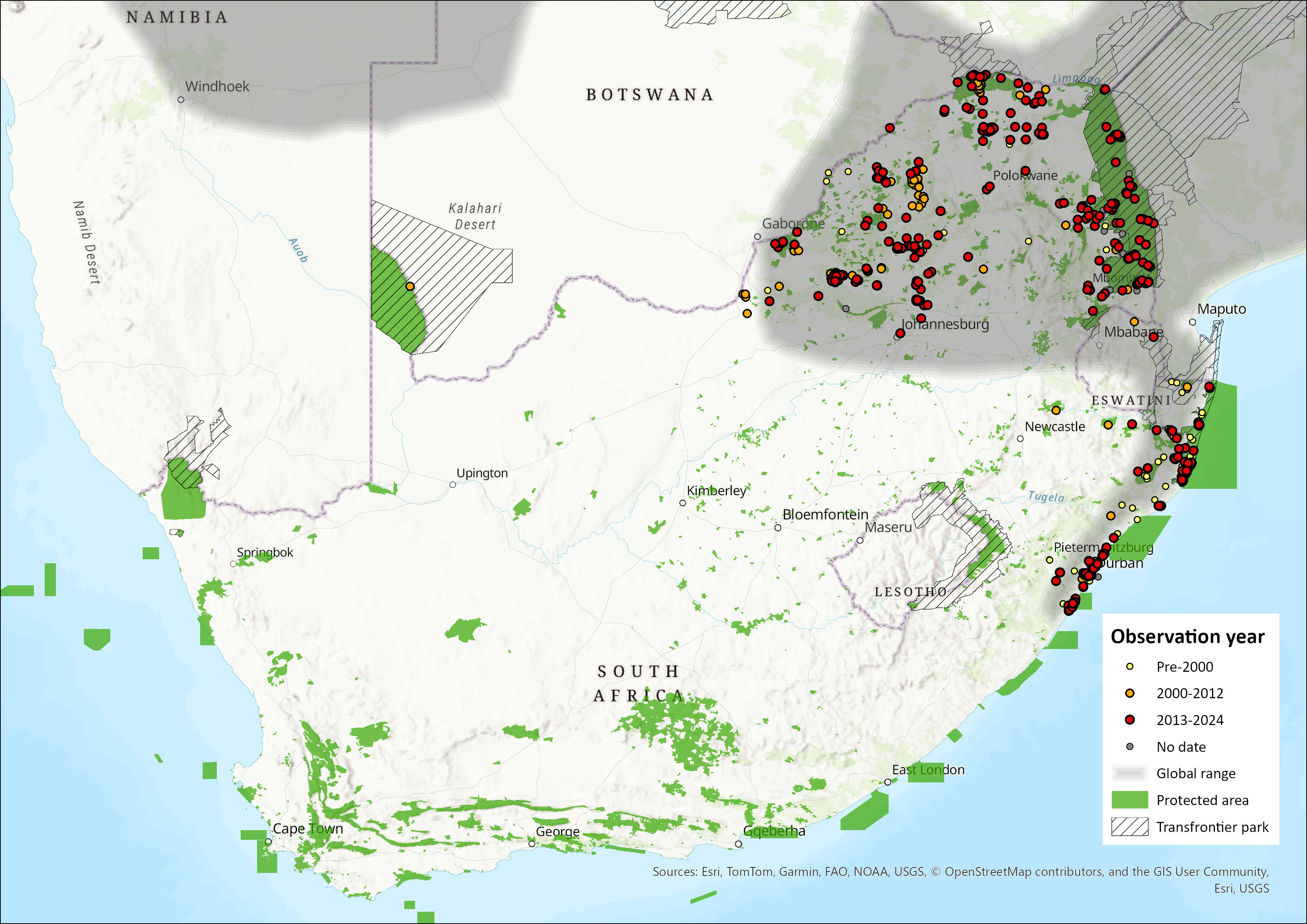
Figure 1. Distribution records for Banded Mongoose (Mungos mungo) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | Resident |
| Benin | Presence Uncertain | Native | – | Resident |
| Botswana | Extant | Native | – | Resident |
| Burkina Faso | Extant | Native | – | Resident |
| Burundi | Extant | Native | – | Resident |
| Cameroon | Extant | Native | – | Resident |
| Central African Republic | Extant | Native | – | Resident |
| Chad | Extant | Native | – | Resident |
| Congo | Extant | Native | – | Resident |
| Congo, The Democratic Republic of the | Extant | Native | – | Resident |
| Côte d’Ivoire | Extant | Native | – | Resident |
| Djibouti | Presence Uncertain | Native | – | Resident |
| Eritrea | Extant | Native | – | Resident |
| Eswatini | Extant | Native | – | Resident |
| Ethiopia | Extant | Native | – | Resident |
| Gabon | Extant | Native | – | Resident |
| Gambia | Extant | Native | – | Resident |
| Ghana | Presence Uncertain | Native | – | Resident |
| Guinea | Extant | Native | – | Resident |
| Guinea-Bissau | Extant | Native | – | Resident |
| Kenya | Extant | Native | – | Resident |
| Malawi | Extant | Native | – | Resident |
| Mali | Extant | Native | – | Resident |
| Mozambique | Extant | Native | – | Resident |
| Namibia | Extant | Native | – | Resident |
| Nigeria | Extant | Native | – | Resident |
| Rwanda | Extant | Native | – | Resident |
| Senegal | Extant | Native | – | Resident |
| Somalia | Extant | Native | – | Resident |
| South Africa | Extant | Native | – | Resident |
| Sudan | Extant | Native | – | Resident |
| Tanzania, United Republic of | Extant | Native | – | Resident |
| Togo | Presence Uncertain | Native | – | Resident |
| Uganda | Extant | Native | – | Resident |
| Zambia | Extant | Native | – | Resident |
| Zimbabwe | Extant | Native | – | Resident |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
It is important to consider a species’ ecological and biological traits to determine the likely mechanisms of climate change impact and quantify these using expert knowledge (Foden & Young 2016). In the North West, the species occurs marginally in the Dry Highveld Grasslands, largely as a result of climate-change directed woody plant establishment in many areas (Power et al. 2019). Food limitations in dry seasons may impact their stress levels and ultimately reproduction (Laver et al. 2020). This may have negative impacts on populations.
Population
Recorded densities vary widely between habitats and locations. In South Africa, Maddock (1988) estimated population density in Vernon Crookes Nature Reserve (KwaZulu-Natal) at 2.4 individuals / km2. Beyond the assessment region, on the Serengeti plains (Tanzania), density was estimated as 2.2 individuals / km2 (Waser et al. 1995). By contrast, a population in Queen Elizabeth National Park (Uganda) was reported to live at higher densities, averaging 18 individuals / km2 (Cant & Gilchrist 2013). It is possible that similar densities may be achieved in the assessment region.
In general, despite them being common in many landscapes, they appear to register low detections on camera traps in various landscapes (Da Rosa 2019; Pretorius 2019; Smith et al. 2023). For instance, Banded Mongoose had low detections by camera traps from the Associated Private Nature Reserve in the Lowveld (Smith et al. 2023), though this may not be reflective of the population status.
In a study in northern KwaZulu-Natal, where a rural pastoral area, mixed farming area and a protected area were compared with camera trapping, the most detections of Banded Mongoose were obtained on farmland, and just less on a protected area, with none detected on rural lands (Da Rosa 2019).
Generation length is estimated to be 4.3 years (Gilchrist & Do Linh San 2016).
Population Information
Current population trend: Unknown, but probably stable based on wide habitat tolerance and lack of threats.
Continuing decline in mature individuals: Unknown, but unlikely.
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of subpopulations: It is not currently possible to determine the extent or number of subpopulations.
Severely fragmented: No. Favourable habitats are relatively well connected across this species’ range.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population Genetics
While no population genetic study has been undertaken on M. mungo within the assessment region, they have been studied in neighbouring Botswana (Verble et al. 2021). Genetic structure was detected between troops, however admixture was also apparent most likely due to high levels of dispersal of individuals into established troops, which could have been facilitated by increased urbanisation (Verble et al. 2021). However, this behaviour is not uniform throughout the species range given that reports from Uganda state dispersal between troops is minimal to absent (Nichols et al. 2012).
Inbreeding depression has been detected within the species despite the species ability to use kin discrimination to minimise inbreeding (Sanderson et al. 2015; Wells 2019). Among troops investigated in Uganda, two thirds were found to be inbred (Schubert et al. 2024).
Given the variable findings of dispersion and levels of inbreeding, it is difficult to extrapolate the genetic structure and health on M. mungo in the assessment region. Consequently, no genetic indicators can be quantified with any level of confidence. It is highly recommended that a population genetic/genomic study be undertaken, specifically looking and the genetic structure, diversity and health of potential subpopulations across the assessment region.
Habitats and ecology
Banded Mongooses occur in a wide range of habitats within savannah and woodland, usually close to water, and are absent from desert, semi-desert and montane regions (Cant & Gilchrist 2013). They are often found in habitats containing termitaria, which are used as den sites with an average den density of 0.71 dens / ha recorded on a mixed cattle and game farm in KwaZulu-Natal (Hiscocks & Perrin 1991a). Banded mongoose used dens for single-night stays, while some dens were used more than once, and they may share dens with Dwarf Mongoose (Helogale parvula), but not simultaneously (Hiscocks and Perrin 1991).
Banded Mongoose have also been observed in towns and villages (Gilchrist et al. 2016). Where Banded Mongoose are in association with humans, they may use anthropogenic structures to den in (Laver 2013; Verble 2019). In KwaZulu-Natal, Banded Mongoose presence has been associated with presence of cattle (Da Rosa 2019), likely related to associated insect abundance with their faeces.
Their diet consists mainly of insects, with other invertebrates, vertebrates (including reptiles, amphibians, the eggs and young of birds, small mammals), and wild fruits also consumed (Hiscocks & Perrin 1991b; Gilchrist et al. 2009; Maddock et al. 2016). Banded Mongooses have been observed to remove ectoparasites (ticks) from Common Warthog (Phacochoerus aethiopicus; Plumptre 2016). They are also known to forage on human garbage (Gilchrist & Otali 2002; Otali & Gilchrist 2004; Laver 2013; Fairbanks Flint et al. 2016).
The Banded Mongoose is a highly social (Schneider & Kappeler 2014) and territorial species that lives in groups of 4–29 individuals with low reproductive skew, i.e. most females breed (Gilchrist et al. 2009), hence making populations less vulnerable to stochastic effects than other social mongoose species such as Suricate (Suricata suricatta) and Common Dwarf Mongoose. Various ecological factors contribute to group living in Banded Mongoose (Gusset 2007).
Home range size is likely larger in more arid areas of South Africa compared to equatorial Uganda (0.61 to 2.01 km2; Gilchrist & Otali 2002). Banded Mongoose troops in northern Botswana have been shown to have home-ranges ranging between 0.39-1.34 km2 (Laver 2013). These may also be similar within the assessment region.
The species is diurnal and foraging distance ranges from 2 to 10 km per day (Neal 1970; Rood 1975, 1986). Dispersal occurs via voluntary fission and eviction (Cant et al. 2013). Within groups, relatedness is high within (but not between) females and males (Cant et al. 2013). In Uganda’s Queen Elizabeth National Park, Banded Mongooses breed up to four times a year, while only one to two litter(s) per year have been recorded in drier regions (Cant & Gilchrist 2013), which may be applicable to much of semi-arid South Africa. Mean age of first conception is 321 days and mean litter size per female at birth (all females) is estimated at 3.32 (Gilchrist et al. 2004), with a gestation period of 90 days (Cant 2000). Within groups, parturition is usually synchronous (Hodge et al. 2011).
Group demography impacts female reproductive success via abortion, eviction and infanticide with younger females bearing the costs (Gilchrist 2006a; Cant et al. 2013). Fecundity and reproductive success are correlated with female age and size (Gilchrist 2006b; Nichols et al. 2012). Survival rate is low in pups (0.299) and high in adults (0.857) (Otali & Gilchrist 2004). Maximum lifespan is 13 years in males and 11 years in females (Cant & Gilchrist 2013).
The species is a carrier of Leptospira interrogans, a pathogen capable of infecting humans (Jobbins et al. 2013), as well as a possible vector of rabies. The Banded Mongoose is susceptible to human tuberculosis (Mycobacterium tuberculosis; Alexander et al. 2002) and the novel derivative M. mungi (Alexander et al. 2010). The latter has shown to be acute and cause high mortality, and to be associated with increased aggression and injury at garbage sites (Fairbanks Flint et al. 2016).
Ecosystem and cultural services: Mongooses in general are known to predate snakes and rats. Banded Mongooses are no exception and will occasionally take both (Gilchrist et al. 2016).
In KwaZulu-Natal, Banded Mongoose presence has been associated with rodent abundance (Da Rosa 2019), so there may be a possibility of an agro-ecosystem service they can offer (Williams et al. 2018), but more likely with insect control
Banded Mongoose may impart some importance to an agro-ecosystems, though it is not always appreciated, however, Limpopo communities even believe them to have a negative impact on themselves (Williams et al. 2018), so these prospects are limiting.
Videographic imagery of Banded Mongoose grooming a Warthog (Phacochoerus africanus) (see Plumptre 2016), has endeared the species a great deal, and popularised them.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
Life History
Generation Length: Estimated to be 4.3 years (Gilchrist & Do Linh San 2016)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
This species is not known to be used or traded in any great deal in the assessment region (Gilchrist et al. 2016).
Consumption of Banded Mongoose meat has been recorded in Botswana (Jobbins et al. 2013) and Mozambique (Fusari & Carpaneto 2006), but is not known within the assessment region (Gilchrist et al. 2016). Banded mongoose body parts have been found at the Faraday market in Gauteng (Williams & Whiting 2016).
The species is not as sought after as the large carnivores, but they are endearing especially where they forage in rest camps, and are certainly appreciated. The species is not reported to be use for sport hunting.
Wildlife ranching may have a positive effect on this species by conserving more suitable habitat (e.g. Cousins et al. 2008, with research in southern Africa suggesting that intensive livestock farming can degrade natural habitat, e.g. Dougill et al. 2006) and possibly helping to connect subpopulations. Of late, a mixed livestock farm with some game, exhibited higher detections of Banded Mongoose than a nearby protected area (Da Rosa 2019), suggesting the importance of some landscapes, although this would be incidental to any use of the species directly.
Local Livelihood: (Not specified)
National Commercial Value: (Not specified)
International Commercial Value: (Not specified)
End Use: (Not specified)
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
There are no major threats to this species (see Gilchrist et al 2016). Diseases and roadkill are perhaps the most prominent.
Roadkill
In northern Limpopo, Collinson et al. (2015) drove a total of 14,400 km over 120 days spanning 10 months between 2011 and 2012, on a 120 km circuit around the Venetia Limpopo Nature Reserve, and a stretch that abutted the Mapungubwe National Park. On this circuit, only one Banded Mongoose was reported to have been killed on these roads (Collinson et al. 2015). Given the 10-month study period, this would amount to 1.2 Mongoose killed by roads per annum.
In the North West, between April 2023 to February 2024, while driving 3005 km in the Bushveld ecoregion (where the species predominates), three road kills of the species were recorded, two of which comprised the same event (NWPG 2024).
Given their appearance on roadside signage to limit roadkill mortality, there may be local threats to some populations even in protected areas (see photo below).

Roadkill Hotspot in the Kruger National Park, as identified from an EWT led project raising awareness of this potential threat (John Power).
Diseases
Banded Mongooses can be susceptible to infectious disease, including human pathogens (Alexander et al. 2002, 2010; Pesapane 2011; Brüns et al. 2017; Verble et al. 2021).
Disease epidemiology has been studied in northern Botswana (Pespane 2011; Fairbanks et al. 2011; Verble et al. 2021). Human fecal waste contamination is an important mechanism for the transmission of pathogens to both humans and Banded Mongoose and is an emerging health threat (Pesapane 2011).
In Banded Mongoose there is no avoidance of clinically diseased mongooses by healthy individuals (Fairbanks et al. 2015). Infected Banded Mongoose may disperse and thus promote disease transmission (Verble 2019), and the risk of pathogens is also spread (Verble et al. 2021).
In 2012 in the Skukuza area of the Kruger National Park, two Banded Mongooses (of 76) were positive for Mycobacterium bovis (BtB) (Brüns et al. 2017).
Sources of infection are likely via scavenging or other spill-over hosts, though it would be important to identify this definitively (Brüns et al. 2017). Given that none had died, the significance is assumed to be low.
Habitat change
Like small mammals, Banded Mongooses may be affected by controlled burning via changes to habitat structure and therefore food availability and predation risk. Research on small mammals has shown that the population effect of fire can be negative or positive (it is species specific; Yarnell et al. 2007). Mongooses may escape fire by using their subterranean dens or termitaria (as for the Short-snouted Elephant Shrew Elephantulus brachyrhynchus; Yarnell et al. 2008) and then may benefit from increased invertebrate availability, firstly via the burn, and subsequently via the fresh growth attracting insects. It is notable that data on fire impact on E. brachyrhynchus and the Lesser Red Musk Shrew (Crocidura hirta), two insectivorous small mammals, indicate no significant impact of controlled burns on survival (Yarnell et al. 2007, 2008). Fire impact on Banded Mongoose has not yet been quantified, but it can be inferred based on their behavioural ecology and potential prey species (Gilchrist et al. 2016).
Persecution – direct and indirectly
Wildlife ranchers do not persecute Banded Mongooses directly, but some animals may be killed as bycatch in control programmes of damage-causing animals (DCAs), especially where poison baits are used. Impact, however, is likely minimal (Gilchrist et al. 2016). For instance, in the North West, since the last assessment (2016), the Problem Animal Control register shows only one complaint or issue received related to the species (North West Directorate of Biodiversity Management records, 2016-2023), so it infrequently causes conflicts with humans, and the status quo remains. In Limpopo rural areas, they are suspected to have a negative impact on livelihoods, but are not known to be persecuted though (Williams et al. 2018).
Body parts use trade
This is perhaps an emerging threat, given the species is recorded in markets (Williams and Whiting 2016), and especially since it was not mentioned in the previous assessment (Gilchrist et al. 2016). However, there is no indication that it is rampant in any way.
Conservation
The Banded Mongoose has been recorded in many national parks and provincial and private nature reserves, as well as on game ranches in five of the nine South African provinces, and occurs in proximity to villages and towns. The most important conservation area is undoubtedly the Kruger National Park, and the adjacent private and state-owned conservation areas on its borders. Other important conservation areas are the Isimangaliso Wetland Park, Pilanesberg Nature Reserve and Marakele National Park.
Recommendations for land managers and practitioners:
- Minimise use of non-selective control methods (e.g. poison) for DCAs.
- Private landowners should ensure that they do not burn the land too frequently and that termite mounds are conserved.
- Create conservancies to protect and connect favourable habitat.
- Limit exposure to human pathogens, including TB, e.g. by restricting mongoose access to garbage pits and human excrement.
Research priorities:
This is one of the few African small carnivore species which has been relatively well studied (see review in Cant & Gilchrist 2013). However, the majority of behavioural ecology research is derived from Uganda (Cant 2000; Gilchrist and Otali 2003; Gilchrist et al. 2004; Otali and Gilchrist 2004; Gilchrist 2006a; b; Hodge et al. 2011; Cant et al. 2013), with some disease monitoring from Botswana (Jobbins et al. 2013; Laver 2013; Fairbanks Flint et al. 2016; Verble et al. 2021). The following research topics will assist in gathering conservation-relevant information:
- Long-term monitoring of (some) subpopulations, particularly those in South Africa and Swaziland
- Evaluation of relative impact of wildlife ranching on habitat and populations.
- Documenting the degree to which controlled burning impacts on population levels.
- Disease evaluation in southern African populations
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas. As confusion with Suricates (and other mongoose species) is possible, a photograph is required for confirmation of identification, especially when sightings are made in areas where the distribution ranges of both mongoose species overlap.
- Limit Banded Mongoose access to garbage pits and human excrement by controlling such access via exclusion if need be (applicable to lodges in game reserves).
Bibliography
Alexander, K.A, Laver, P.N., Michel, A.L., Pleydell, E., Williams, M.C., van Helden, P.D., Warren, R.M. and Gey van Pittius, N.C. 2010. Novel Mycobacterium tuberculosis complex pathogen, M. mungi. Emerging Infectious Diseases 16: 1296–1299.
Alexander, K.A., Pleydell, E., Williams, M., Lane, E.P., Myange, J.F.C. and Michael, A.F. 2002. Mycobacterium tuberculosis: an emerging disease of free-ranging wildlife. Emerging Infectious Diseases 8: 598–601.
Brüns, A.C., Tanner, M., Williams, M.C., Botha, L., O’Brien, A., Fosgate, G.T., Van Helden, P.D., Clarke, J. and Michel, A.L. 2017. Diagnosis and implications of Mycobacterium bovis infection in banded mongooses (Mungos mungo) in the Kruger National Park, South Africa. Journal of wildlife diseases. 53(1):19-29.
Cant, M.A. 2000. Social control of reproduction in banded mongooses. Animal Behaviour 59: 147-158.
Cant, M.A. and Gilchrist, J.S. 2013. Mungos mungo Banded Mongoose. In: J. Kingdon and M. Hoffmann (eds), The Mammals of Africa. V. Carnivores, Pangolins, Equids and Rhinoceroses, pp. 354-360. Bloomsbury, London, UK.
Cant, M.A., Vitikainen, E. and Nichols, H.J. 2013. Demography and social evolution of banded mongooses. Advances in the Study of Animal Behaviour 45: 407-445.
Collinson, W.J., Reilly, B.K., Parker, D.M., Bernard, R.T. and Davies-Mostert, H.T. 2015. An inventory of vertebrate roadkill in the greater Mapungubwe Transfrontier conservation area, South Africa. African Journal of Wildlife Research 45(3): 301-311.
Cousins, J.A., Sadler, J.P. and Evans, J. 2008. Exploring the role of private wildlife ranching as a conservation tool in South Africa: stakeholder perspectives. Ecology and Society 13: 43.
Da Rosa, B.R.P. 2019. Optimizing Small Mammal Relative Abundance Measures Using Non-Invasive Sampling and Assessment of its Contribution to Occupancy Modelling of Small Carnivores in Dry Woodland Savannah of South Africa. Master’s thesis, University of Lisbon, Lisbon.
Dougill, A.J., Akanyang, L., Perkins, J.S., Eckardt, F.D., Stringer, L.C., Favretto, N., Atlhopheng, J. and Mulale, K. 2016. Land use, rangeland degradation and ecological changes in the southern Kalahari, Botswana. African Journal of Ecology 54: 59–67.
Fairbanks, B.M., Hawley, D.M. and Alexander, K.A. 2015. No evidence for avoidance of visibly diseased conspecifics in the highly social banded mongoose (Mungos mungo). Behavioral Ecology and Sociobiology. 69:371-381.
Fairbanks Flint, B., Hawley, D.M. and Alexander, K.A. 2016. Do not feed the wildlife: associations between garbage use, aggression, and disease in banded mongooses (Mungos mungo). Ecology & Evolution 6: 5932–5939.
Foden, W.B. and Young, B.E. 2016. IUCN SSC Guidelines for Assessing Species’ Vulnerability to Climate Change. Version 1.0. Occasional Paper of the IUCN Species Survival Commission No. 59. Cambridge, UK and Gland, Switzerland: IUCN Species Survival Commission. x+114pp.
Fusari, A. and Carpaneto, G.M. 2006. Subsistence hunting and conservation issues in the game reserve of Gile, Mozambique. Biodiversity and Conservation 15(8): 2477-2495.
Gilchrist JS, Do Linh San E. 2016. Mungos mungo. The IUCN Red List of Threatened Species 2016.
Gilchrist, J.S. 2006a. Female eviction, abortion, and infanticide in banded mongooses (Mungos mungo): implications for social control of reproduction and synchronized parturition. Behavioral Ecology 17: 664-669.
Gilchrist, J.S. 2006b. Reproductive success in a low skew, communal breeding mammal: the banded mongoose, Mungos mungo. Behavioral Ecology and Sociobiology 60: 854-863.
Gilchrist, J.S. and Otali, E. 2002. The effects of refuse-feeding on home-range use, group size, and intergroup encounters in the banded mongoose. Canadian Journal of Zoology 80: 1795-1802.
Gilchrist, J.S., Jennings, A.P., Veron, G. and Cavallini, P. 2009. Family Herpestidae (Mongooses). In: D.E. Wilson and R.A. Mittermeier (eds), Handbook of the Mammals of the World. 1. Carnivores, pp. 262-328. Lynx Edicions, Barcelona, Spain.
Gilchrist, J.S., Otali, E. and Mwanguhya, F. 2004. Why breed communally? Factors affecting fecundity in a communal breeding mammal: the banded mongoose (Mungos mungo). Behavioral Ecology and Sociobiology 57: 119-131.
Gilchrist, J.S., Stuart, C., Stuart, M. and Do Linh San, E. 2016. A conservation assessment of Mungos mungo. In Child MF, Roxburgh L, Do Linh San E, Raimondo D, Davies-Mostert HT, editors. The Red List of Mammals of South Africa, Swaziland and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Gusset, M. 2007. Banded together: a review of the factors favouring group living in a social carnivore, the banded mongoose Mungos mungo (Carnivora: Herpestidae). Mammalia. 71(1-2):80-82.
Hiscocks, K. and Perrin, M.R. 1991a. Den selection and use by dwarf mongooses and banded mongooses in South Africa. South African Journal of Wildlife Research 21: 119-122.
Hiscocks, K. and Perrin, M.R. 1991b. A dietary comparison between two sympatric vierrids, Helogale parvula (Sundevall 1846) and Mungos mungo (Gmelin 1788). Journal of African Zoology 105: 307–312.
Hodge, S.J., Bell, M.B.V. and Cant, M.A. 2011. Reproductive competition and the evolution of extreme birth synchrony in a cooperative mammal. Biology Letters 7: 54-56.
Jobbins, S.E., Sanderson, C.E. and Alexander, K.E. 2013. Leptospira interrogans at the human–wildlife interface in northern Botswana: a newly identified public health threat. Zoonoses and Public Health 61: 113-123.
Kingdon, J. 1997. The Kingdon Field Guide to African Mammals (first edition}. Academic Press, San Diego, California, USA.
Laver, P.N. 2013. The foraging ecology of banded mongooses (Mungos mungo): epidemiological and human-wildlife conflict implications. Doctoral dissertation, Virginia Polytechnic Institute and State University, USA.
Laver PN, Ganswindt A, Ganswindt SB, Alexander KA. 2020. Effect of food limitation and reproductive activity on fecal glucocorticoid metabolite levels in banded mongooses. BMC ecology 20:1-24.
Maddock, A.H, Do Linh San, E. and Perrin, M.R. 2016. Some data on the feeding habits of the banded mongoose in a coastal area (South Africa). African Journal of Ecology 54: 245–247.
Maddock, A.H. 1988. Resource partitioning in a viverrid assemblage. Ph.D. Thesis, University of Natal.
Monadjem, A. 1998. The mammals of Swaziland. Conservation Trust of Swaziland and Big Games Parks, Mbabane, Swaziland.
Neal, E. 1970. The banded mongoose Mungos mungo Gmelin. East African Wildlife Journal 8: 53-71.
NWPG (North West Provincial Government). 2024. The impact of roadkill on mammal populations in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Otali, E. and Gilchrist, J.S. 2004. The effects of refuse feeding on body condition, reproduction and survival of banded mongooses. Journal of Mammalogy 85: 491-497.
Pesapane, R.R. 2011. Tracking Pathogen Transmission at the Human-Wildlife Interface: Banded Mongoose (Mungos mungo) and Escherichia coli as a Model System in Chobe, Botswana. Msc thesis, Virginia Tech, Blacksburg.
Plumptre, A. 2016. Banded mongooses grooming warthogs. Suiform Soundings 14: 31.
Power. R.J., Van Straaten, A., Schaller, R., Mooke, M., Boshoff, T. and Nel, H.P. 2019. An inventory of mammals of the North West Province, South Africa. Annals of the Ditsong Museum of Natural History 8: 6-29.
Pretorius, M. 2019. Mesocarnivores in Protected Areas: ecological and anthropogenic determinants of habitat use in northern Kwa-Zulu Natal, South Africa. MSc thesis, University of Cape Town, Cape Town.
Rood, J.P. 1975. Population dynamics and food habits of the banded mongoose. East African Wildlife Journal 13: 89-111.
Rood, J.P. 1986. Ecology and social evolution in the mongooses. In: D. Rubenstein and R. Wrangham (eds), Ecological Aspects of Social Evolution, pp. 131-152. Princeton University Press, Princeton, USA.
Sanderson, J.L., Wang, J., Vitikainen, E.I., Cant, M.A. and Nichols, H.J. 2015. Banded mongooses avoid inbreeding when mating with members of the same natal group. Molecular Ecology 24(14):3738-51.
Schneider, T.C. and Kappeler, P.M. 2014. Social systems and life‐history characteristics of mongooses. Biological Reviews 89(1):173-198.
Schubert, N., Stober, C., Born, M., Mwanguhya, F., Businge, R., Kyambulima, S., Mwesige, K., Cant, M.A., Nichols, H.J. and Winternitz, J.C. 2024. Banded mongooses discriminate relatedness and MHC diversity in unfamiliar conspecifics. bioRxiv, pp.2024-12.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Smith, K., Venter, J.A., Peel, M., Keith, M. and Somers, M.J. 2023. Temporal partitioning and the potential for avoidance behaviour within South African carnivore communities. Ecology and Evolution. 13(8):e10380.
Van Rompaey, H. and Sillero-Zubiri, C. 2013. Mungos gambianus Gambian Mongoose. In: J. Kingdon and M. Hoffmann (eds), The Mammals of Africa. V. Carnivores, Pangolins, Equids and Rhinoceroses, pp. 353-354. Bloomsbury, London, UK.
Verble, K.M. 2019. Understanding the Influence of Banded Mongoose (Mungos mungo) Social Structuring on Disease Transmission Using Molecular Tools. Msc dissertation, Virginia Tech, USA.
Verble, K., Hallerman, E.M. and Alexander, K.A. 2021. Urban landscapes increase dispersal, gene flow, and pathogen transmission potential in banded mongoose (Mungos mungo) in northern Botswana. Ecology and Evolution. 11(14):9227-9240.
Veron, G., Colyn, M., Dunham, A.E., Taylor, P. and Gaubert, P. 2004. Molecular systematics and origin of sociality in mongooses (Herpestidae, Carnivora). Molecular phylogenetics and evolution. 30(3):582-98.
Veron, G., Patou, M.L. and Jennings, A.P. 2022. Systematics and evolution of the mongooses (Herpestidae, Carnivora). In Small Carnivores: Evolution, Ecology, Behaviour, and Conservation. Do Linh San, E., Sato, J. J., Belant, J. L. and Somers, M. J. (Eds). Pages 61-78.
Waser, P.M., Elliott, L.F., Creel, N.M. and Creel, S.R. 1995. Habitat variation and mongoose demography. In: A.R.E. Sinclair and P. Arcese (eds), Serengeti II: dynamics, management, and conservation of an ecosystem, pp. 421-447. University of Chicago Press, Chicago, IL, USA.
Wells, D.A. 2019. Causes and consequences of inbreeding in the banded mongoose. Liverpool John Moores University (United Kingdom).
Williams, V.L. and Whiting, M.J. 2016. A picture of health? Animal use and the Faraday traditional medicine market, South Africa. Journal of Ethnopharmacology. 179:265-273.
Yalden, D.W., Largen, M.J., Kock, D. and Hillman, J.C. 1996. Catalogue of the mammals of Ethiopia and Eritrea. 7. Revised checklist, zoogeography and conservation. Tropical Zoology 9: 73-164.
Yarnell, R.W., Scott, D.M., Chimimba, C.T. and Metcalfe, C.J. 2007. Untangling the roles of fire, grazing and rainfall on small mammal communities in grassland ecosystems. Oecologia 154: 387–402.
Yarnell, R.W., Metcalfe, D.J., Dunstone, N., Burnside, N. and Scott, D.M. 2008. The impact of fire on habitat use by the short-snouted elephant shrew (Elephantulus brachyrhynchus) in North West Province, South Africa. African Zoology 43: 45-52.