

African Savanna Hare
Lepus victoriae

2025 Red list status
Regional Population Trend
Declining
Change compared
to 2016
No Change
Overview
Lepus victoriae – Thomas, 1893
ANIMALIA – CHORDATA – MAMMALIA – LAGOMORPHA – LEPORIDAE – Lepus – victoriae
Common Names: African Savanna Hare (English), Afrikanischer Savannen Hase (German), Lievre Des Savanes (French)
Synonyms: Lepus crawshayi de Winton, 1899; Lepus microtis Heuglin, 1865, L. whytei. Fifteen listed in total (Happold 2013b).
Taxonomic Note: Previously this was assessed as Lepus microtis. Here, L. victoriae is maintained as a valid species following Happold (2013).
The taxonomic status of this species and synonyms (including microtis, crawshayi, canopus, whytei and zechi) are uncertain (Happold 2013, Awadi and Ben Slimen 2018). Hoffmann and Smith (2005) recognised four subspecies: Lepus microtis angolensis, L. m. microtis, L. m. senegalensis, and L. m. whytei. Happold (2013) lists a total of 15 synonyms. This species was formerly included in L. saxatilis and has been classified under several different names (saxatilis, crawshayi, whytei, and victoriae) (Hoffmann and Smith 2005). Ben Slimen et al. (2008) suggest that the name Lepus victoriae be retained and microtis considered a “nomen dubium” referencing Petter (1959) and Angermann (1965).
In Africa, L. capensis sensu lato, in addition to five other Lepus species are currently classified (Lepus saxatilis, Lepus victoriae, Lepus habessinicus, Lepus fagani and Lepus starcki), primarily distinguished by morphological characters (Happold 2013). However, their evolutionary history, genetic differentiation and distribution remain poorly understood. This is confounded by the frequent sharing of mitochondrial DNA (mtDNA) between species and discordant patterns between mtDNA and with nuclear DNA (Ben Slimen et al.)
Red List Status: Least concern
Assessment Information
Assessors: Wilson-Hartmann, B.1 & da Silva, J.2
Reviewer: Moodley, S.3
Institutions: 1McGregor Museum, Kimberley, 2South African National Biodiversity Institute, 3Private
Previous Assessors & Reviewers: Robinson, T.J., Relton, C. & Child, M.F.
Previous Contributors: Roxburgh, L. & Smith, A.
Assessment Rationale
Lepus victoriae in South Africa occurs widely within the assessment region in many habitats both protected and transformed. Ongoing molecular research is urgently required to resolve their taxonomic statuses and relationships to delimit their geographical ranges, and thus assess threat severity, more accurately. These species may require reassessment when such data becomes available.
Although local declines may be occurring due to overhunting for bushmeat and fur in some areas, there was no evidence to suggest a significant widespread population decline during past assessments However, with the outbreak of Rabbit Haemorrhagic Disease Virus strain 2 (RHDV-2) in November 2022 in the Northern Cape Province, South Africa that subsequently spread to all the provinces (DALRRD 2024), urgent investigations are needed to ascertain the effect on populations and subpopulations. It is relatively abundant in the northeastern regions of the assessment region. Thus, the Least Concern listing is retained until taxonomic resolution and population data suggest otherwise.
Regional population effects: Taxonomic revision is required to substantiate the geographical range limits of Lepus species in southern Africa.
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended citation: Wilson-Hartmann B & da Silva JM. 2025. A conservation assessment of Lepus victoriae. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
Lepus victoriae has an extensive African distribution, which extends along the Atlantic coast (Guinea, The Gambia, Senegal, Mauritania and Western Sahara), across the Sahel and into western Kenya and Ethiopia, southwards to South Africa, Mozambique, Zimbabwe, Botswana and Namibia (Hoffmann & Smith 2005; Smith & Johnston 2008). Additionally, an isolated population has been found in western Algeria (Flux & Angermann 1990; Hoffmann & Smith 2005). However, taxonomic uncertainty again limits confidence in geographical distribution with Happold (2013) stating that the species is not found in south-western Africa (being instead replaced by L. saxatilis). It coexists with the Cape Hare (L. capensis) over much of its distribution (northeastern parts of the country, including the North West, Free State, KwaZulu-Natal, Mpumalanga, Gauteng and Limpopo provinces, as well as the easterly regions of the Eastern Cape Province); they are separated largely by habitat with L. victoriae preferring mountain regions (Wilson et al. 2016).
Taxonomic resolution and vetting of museum records through molecular research is needed urgently to resolve the tentative geographical ranges.
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): (Not specified)
Elevation Upper Limit (in metres above sea level): 3 600m
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
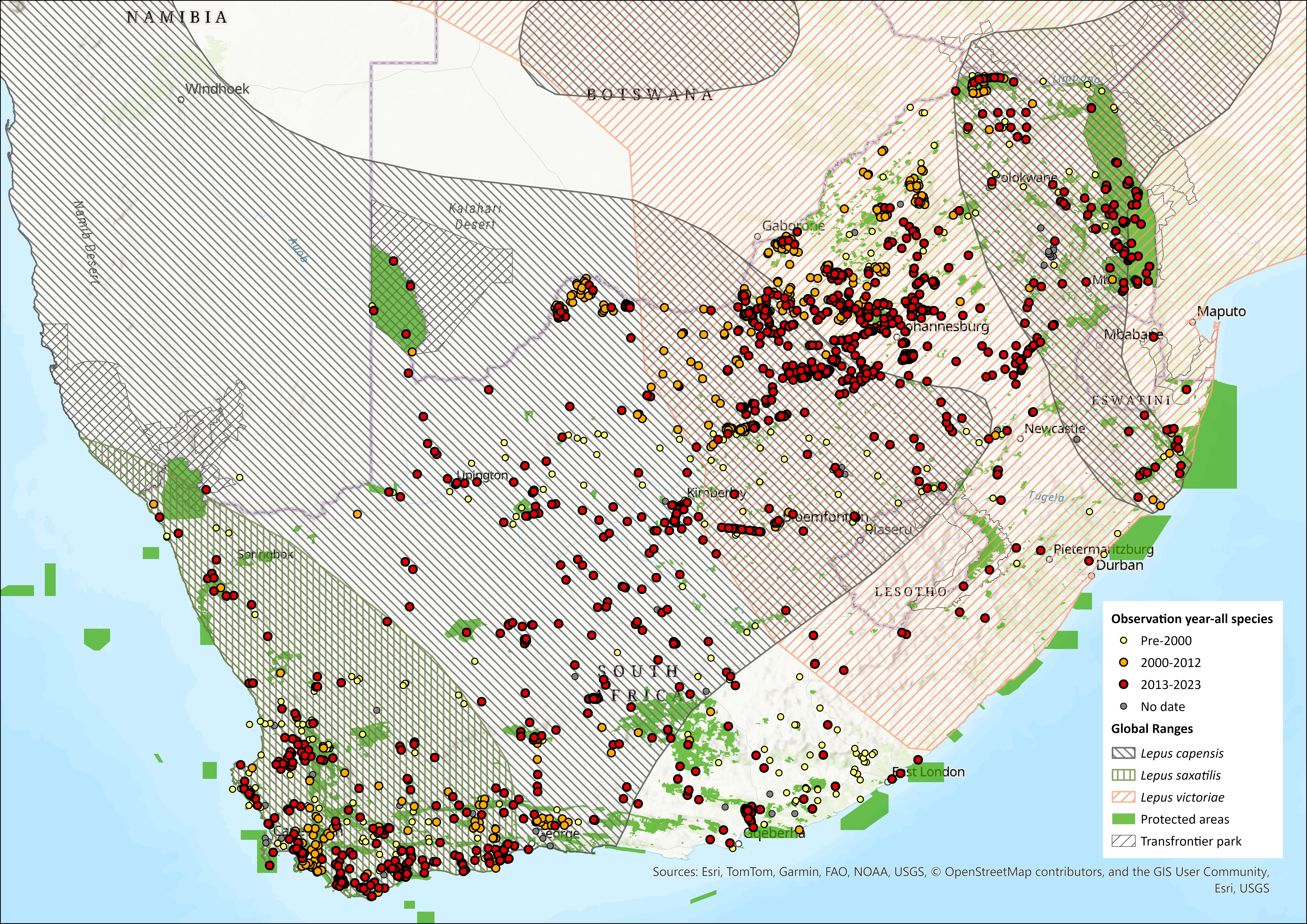
Figure 1. Distribution records for African Savanna Hare (Lepus victoriae) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Algeria | Extant | Native | – | – |
| Angola | Extant | Native | – | – |
| Benin | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Burkina Faso | Extant | Native | – | – |
| Burundi | Extant | Native | – | – |
| Cameroon | Extant | Native | – | – |
| Central African Republic | Extant | Native | – | – |
| Chad | Extant | Native | – | – |
| Congo, The Democratic Republic of the | Extant | Native | – | – |
| Côte d’Ivoire | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Ethiopia | Extant | Native | – | – |
| Gambia | Extant | Native | – | – |
| Ghana | Extant | Native | – | – |
| Guinea | Extant | Native | – | – |
| Guinea-Bissau | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Lesotho | Extant | Native | – | – |
| Malawi | Extant | Native | – | – |
| Mali | Extant | Native | – | – |
| Mauritania | Extant | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Niger | Extant | Native | – | – |
| Nigeria | Extant | Native | – | – |
| Rwanda | Extant | Native | – | – |
| Senegal | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Togo | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Western Sahara | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
Whilst climate change per se is unlikely to have a dramatic effect on this species during this assessment period, climate change is already affecting the spread of infectious diseases beyond their typical geographic reach. Whilst this may not be the cause of the outbreak of Rabbit Haemorrhagic Disease Virus strain 2 (RHDV-2) in South Africa at the end of 2022, it can assist in the spread of the virus and increase the susceptibility of hares and rabbits to the effects of droughts and periods of flooding. Often, the cross-border spread of infectious diseases is further exacerbated by the lack of global governance (that led to the introduction of this disease into South Africa and allowed it to spread to neighbouring countries), policies or a consensus to mitigate climate change. As a result, the current and future burden on humans, animals and plants is significant, especially if these infectious diseases cause large scale outbreaks such as RHDV-2.
Population information
No detailed population or density estimates are available for L. victoriae but it is considered a successful and common species (Flux & Angermann 1990; Smith & Johnston 2008).
Lepus species overall generally exhibit a fast population turnover and a high rate of reproduction (Kryger et al. 2004b). While a slow decline has been predicted due to habitat loss and hunting pressure (Kryger et al. 2004a), it is expected that land conversion from livestock to wildlife may benefit Lepus species and counteract population declines by creating and reconnecting suitable habitat patches. However, the status of scattered subpopulations and isolated subspecies is largely unknown and may be severely threatened by the recent RHDV-2 outbreak that is affecting all Lepus and Pronolagus populations throughout southern Africa.
Population Information
Continuing decline in mature individuals? Yes
Extreme fluctuations in the number of subpopulations: No
Continuing decline in number of subpopulations: Yes
All individuals in one subpopulation: No
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Unknown
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: Unknown
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: Unknown
Probability of extinction in the wild within 100 years: Unknown
Population genetics
While molecular studies have been conducted on the genus Lepus, and in particular L. capensis (e.g., Suchentrunk et al. 2009; Lado et al. 2019), little molecular research has investigated L. victoriae in detail.
While few records are noted for L. victoriae, it is believed to be widely distributed across the northeastern parts of South Africa. Given the ability of Lepus species to utilise modified habitats and be able to disperse, the species is believed to exist as a metapopulation. Assuming no obstructions to gene flow and no known subpopulation extinctions, the Convention on Biological Diversity’s Global Biodiversity Framework’s (GBF’s) complementary genetic indicator – proportion of populations maintained (PM) – would receive a value of 1.0 (all populations remaining).
While no census size (Nc) data exists and there is a lack of general density estimates, the assumption of no population structure within the assessment region, the likely presence of gene flow between neighbouring countries, and a rapid turnover in offspring, it is very likely the effective population size (Ne) of the species in the region exceeds the Ne 500, suggesting a healthy and stable population. Based on these assumptions, the GBF’s headline genetic indicator – proportion of populations with an effective population size, Ne, greater than 500 – would receive a value of 1.0 (1 metapopulation/1 with Ne 500 = 1.0).
Given the limited demographic and genetic data available for this species, this information should be taken with caution. It is recommended that a population genetic study be undertaken across the species range within the assessment region and beyond to investigate the genetic structure and diversity in L. victoriae.
Habitats and ecology
Little data is currently available regarding the general ecology of L. victoriae (Boitani et al. 1999). The African Savanna Hare occurs in scrub, bush, and grassland habitats, preferring scrubby and montane habitats. It has been reported in mountain regions up to ¢.3 600 m on Mount Kenya and is also recorded in semiarid grasslands. It coexists with the Cape Hare (L. capensis) over much of its distribution; they are separated largely by habitat. Where L. capensis and L. victoriae co-occur, the latter prefers scrubbier and more montane habitats (Flux & Angermann 1990). Helping it to cut and nip at vegetation are two pairs of chisel-shaped incisors, which display a distinctive groove pattern that sets it apart from L. capensis. L. victoriae hasbeen recorded in grassland associations of Themeda triandra and Imperata cylindrical (both Poaceae) in Ruanda-Urundi and Tarchonanthus camphoratus (Asteraceae) scrub in East Africa (Wilson et al. 2016). It is suspected that this species can thrive in agricultural landscapes.
The diet of this species varies with habitat type, and similar to other Lepus species, females breed throughout the year and mean litter size was 1-6 precocious young Flux and Angermann (1990), and the numbers of litters per year were 6-8, resulting in a mean of 11-2 young/female/year (Wilson et al. 2016).
L. victoriae is strictly nocturnal and spend most of the day lying in a “form”—a depression in the ground or under bushes. Home ranges are 5-10 ha. They live mostly alone but are sometimes seen in groups of 2-3 individuals while feeding.
Ecosystem and cultural services: Lepus species form a valuable prey component for a broad array of carnivores, such as Black-backed Jackals (Lupulella mesomelas); Kamler et al. 2012; Bagniewska & Kamler 2014) and Verreaux’s Eagles (Aquila verreauxii; Cruz-Uribe & Klein 1998). Hares were found to be the dominant prey taxon for Martial Eagles (Polemaetus bellicosus) in the Cape region of South Africa, making up 50% of the total diet (Boshoff et al. 1990). Lepus species were also found to constitute a small component of the diet of Cape Foxes (Vulpes chama) in the Free State (Kamler et al. 2012).
IUCN Habitats Classification Scheme
Life History
Generation Length: Unknown
Age at maturity: female or unspecified: 1 year
Age at Maturity: Male: 1 year
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)1-5 years
Average Reproductive Age: (Not specified)
Maximum Size (in cms): 55cm
Size at Birth (in cms): (Not specified)
Gestation Time: 42 days
Reproductive Periodicity: All months
Average Annual Fecundity or Litter Size: LS=1.6 (average)
Natural Mortality: (Not specified)
Breeding Strategy
Does the species lay eggs?: No
Does the species give birth to live young: Yes
Does the species exhibit parthenogenesis: No
Does the species have a free-living larval stage?: No
Does the species require water for breeding?: No
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
Lepus species are hunted recreationally for sport, bushmeat and fur at a subsistence level in the assessment region. However, this is not expected to have a substantial effect on the population. Both L. capensis and L. saxatilis were listed as species utilised for traditional medicine in South Africa, as they are believed to have medicinal or curative properties (Maliehe 1993; Ntiamoa-Baidu 1997) and this likely to be the same with L. victoriae.
Local Livelihood: Unknown
National Commercial Value: No
International Commercial Value: No
End Use: Unknown
Is there harvest from captive/cultivated sources of this species? No
Harvest Trend Comments: Unknown
Threats
Across their range, as well as within the assessment region, Lepus species are threatened by habitat loss and fragmentation because of urban sprawl, agricultural encroachment, commercial plantations, and infrastructure development for tourism (Drew et al. 2004; Kryger et al. 2004a). Additionally, these species are threatened by hunting pressure through both recreational sport hunting, as well as subsistence hunting for bushmeat and fur (Kryger et al. 2004a). While agricultural and urban expansion may not necessarily cause direct declines, they may increase hunting pressures. Hunting pressure is likely to cause local subpopulation declines. For example, drastic population declines have been observed in KwaZulu-Natal, specifically in Harding and Port Shepstone in southern KwaZulu-Natal (Kryger et al. 2004a). Lepus species may also be vulnerable to predation by domestic dogs associated with farming areas.
However, a more urgent threat has recently emerged. In October 2022, reports of unusual Lepus deaths were received from the Middelpos, Springbok and Fraserburg areas in the Northern Cape. In November 2022, affected carcasses tested by Onderstepoort Veterinary Research Laboratory, the deaths were confirmed to as the result of the highly contagious and acute fatal hepatitis of Leporids called Rabbit Haemorrhagic Disease Virus 2 strain (RHDV-2). The new variant RHDV-2 affects rabbits, but also causes fatal RHD in various Lepus species, including Sardinian Cape hares (L. capensis mediterraneus), Italian hares (L. corsicanus), and mountain hares (L. timidus) (Rocchi & Dagleish 2018). This disease, which originated in China in 1984, primarily affects all rabbit and hares. Caused by a calicivirus, it was previously absent in South Africa. Infection typically occurs by the oral route. It spreads very easily, normally through direct contact between individuals but is also carried by insects, birds, scavengers feeding on carcasses, contaminated food or urine and faeces, and by humans on clothes, shoes, tools and car tyres. The virus can persist in infected meat (even if frozen) and for long periods in decomposing carcasses (Gleeson & Petritz 2020). Incubation varies from one to three days, and death usually occurs 12-36 hours after the onset of fever. The main clinical manifestations of the acute infection are nervous and respiratory signs, apathy and anorexia. In RHDV-2, clinical signs and mortality are observed even in young animals from 7 to 15 days of age onwards. Indirect control of the disease is easily achieved by vaccination. RHDV has never been reported in humans and other mammals. Outbreaks tend to be seasonal (typically peak breeding seasons) in wild rabbit and hare populations where adults have survived infection and are immune (Kerr & Donnelly 2013). As young kits grow up and stop nursing, they no longer receive the antibodies in their mother’s milk and become susceptible to infection. Mortality rates are high and up to 90% of infected individuals die (WOAH 2025). Not all rabbits exposed to RHDV-2 become overtly ill. A small proportion of infected rabbits clears the virus without developing signs of disease (Kerr & Donnelly 2013). Asymptomatic carriers also occur, and can continue to shed virus for months, thereby infecting other animals. Surviving rabbits develop a strong immunity to the specific viral variant with which they were infected (Gleeson & Petritz 2020).
This outbreak is of concern for indigenous rabbit and hare species due to their historic naivety to the virus and thus susceptibility to the virus. Whilst a vaccine is available for captive situations overseas, there is no specific treatment for RHDV-2 and management is limited to preventing further spread.
Often, the cross-border spread of infectious diseases is further exacerbated by the lack of global governance, a factor that led to the introduction of this disease into South Africa and allowed it to spread to neighbouring countries.
Conservation
Lepus species are widely distributed across the assessment region, occurring within numerous protected areas, including both formally and privately protected areas (Kryger et al. 2004a). A call for sustainable utilisation of these species is recommended (Kryger et al. 2004a), as they may constitute a low-carbon source of protein and may economically benefit local communities and landowners (Asibey 1974). The development of conservancies to protect appropriate habitats for local subspecies and forms is recommended. Taxonomic resolution at a species and subspecies level is required for the Lepus genus to ensure that localised or isolated populations and subspecies are conserved. Research into the ecology of L. victoriae is lacking and is restricted to localised regions of its distribution (Boitani et al. 1999), thus further investigations are necessary.
Recommendations for land managers and practitioners:
- This species would benefit from suitable land management: landowners should retain corridors of grassland between grazed areas and decrease stocking rates to avoid overgrazing and the loss of grass cover.
Research priorities:
- Taxonomic revision of the Lepus victoriae, including the identification of isolated populations and localised subspecies.
- Survey studies to effectively document the distributional limits and geographical overlap of species and subspecies.
- Population size and trends.
- Effects of wildlife ranching on Lepus species within the assessment region.
- Determination of the impact of RHDV-2 on the southern African population, particularly at subpopulation level.
Encouraged citizen actions:
- Landowners should create conservancies to utilise this species sustainably.
- Refrain from having too many dogs on farms.
- Report sightings of free-roaming individuals on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas to enhance the distribution maps.
- Report unusual death outbreaks potentially linked to RHDV-2 and prevent the spread of the disease.
Bibliography
Alves, P.C., Harris, D.J., Melo-Ferreira, J., Branco, M., Ferrand, N. et al. 2006. Hares on thin ice: Introgression of mitochondrial DNA in hares and its implications for recent phylogenetic analyses. Molecular Phylogenetics and Evolution 40 (2): 640-641.
Angermann, R. 1965. Revision der Palaarkitschen und Athiopischen Arten der Gattung Lepus (Leporidae, Lagomorpha). Humboldt University of Berlin, Germany.
Asibey EOA. 1974. Wildlife as a source of protein in Africa south of the Sahara. Biological Conservation 6: 32–39.
Bagniewska, J.M, Kamler JF. 2014. Do black-backed jackals affect numbers of smaller carnivores and prey? African Journal of Ecologyvvvv 52: 546–567.
Ben Slimen, H., Suchentrunk, F., Stamatis, C., Mamuris, Z., Sert, H., Alves, P.C., Kryger, U., Shahin, A.B., and Ben Ammar Elgaaied, A. 2008. Population genetics of cape and brown hares (Lepus capensis and L. europaeus): A test of Petter’s hypothesis of conspecificity. Biochemical Systematics and Ecology 36: 22-39.
Ben Slimen, H., Suchentrunk, F., Memmi, A. et al. 2006. Evolutionary relationships among hares from North Africa (Lepus sp. or Lepus spp.), cape hares (L. capensis) from South Africa, and brown hares (L. europaeus), as inferred from mtDNA PCR‐RFLP and allozyme data. Journal of Zoological Systematics and Evolutionary Research 44(1): 88 – 99.
Ben Slimen, H., Suchentrunk, F. and Ben Ammar Elgaaied, A. 2008. On shortcomings of using mtDNA sequence divergence for the systematics of hares (genus Lepus): An example from cape hares. Mamm Biol 73, 25–32.
Boitani, L., Corsi, F., De Biase, A. 1999. A Databank for the Conservation and Management of the African Mammals. Istituto di Ecologia Applicata, Rome, Italy.
Boshoff, A.F., Palmer, N.G., Avery, G. 1990. Regional variation in the diet of martial eagles in the Cape Province, South Africa. African Journal of Wildlife Research 20: 57–68.
Cruz-Uribe, K., Klein, R.G. 1998. Hyrax and hare bones from modern South African eagle roosts and the detection of eagle involvement in fossil bone assemblages. Journal of Archaeological Science 25: 135–147.
Drew, C., Barcelo, I. and Saji, A. 2004. A Report on the Findings of an Ecological and Environmental Survey of the Sweihan Region. Environmental Research and Wildlife Development Agency, Abu Dhabi, UAE.
Ellerman, J.R. and Morrison-Scott, T.C.S. 1951. Checklist of Palaearctic and Indian Mammals 1758 to 1946. British Museum (Natural History), London, UK.
Ellerman, J.R., Morrison-Scott, T.C.S. and Hayman, R.W. 1953. Southern African Mammals 1758 to 1951: A Reclassification. British Museum (Natural History), London, UK.
Flux, J. C. 1981. Reproductive strategies in the genus Lepus. University of Guelph, Guelph.
Flux, J.E.C. and Angermann, R. 1990. The hares and jackrabbits. In: J.A. Chapman and J.E.C. Flux (eds), Rabbits, Hares and Pikas: Status Survey and Conservation Action Plan, pp. 61-94. IUCN, Gland, Switzerland.
Gleeson, M. and Petritz, O.A. 2020. Emerging Infectious Diseases of Rabbits. Veterinary Clinics of North America: Exotic Animal Practice. 23 (2): 249–261.
Happold, D.C.D. 2013. Lepus victoriae African Savanna Hare. In: Happold DCD (ed.), Mammals of Africa Volume III: Rodents, Hares and Rabbits, pp. 706–707. Bloomsbury Publishing, London, UK.
Hoffmann, R.S. and Smith, A.T. 2005. Order Lagomorpha. In: D.E. Wilson and D.M. Reeder (eds), Mammal Species of the World, pp. 185-211. Johns Hopkins University Press, Baltimore, Maryland, USA.
Kamler, J. F., Stenkewitz, U., Klare, U. Jacobsen, N. F., and Macdonald, D. W. 2012b. Resource partitioning among cape foxes, bat-eared foxes, and black-backed jackals in South Africa. Journal of Wildlife Management 76: 1241-1253.
Kerley, G.I.H. 1990. Browsing by Lepus capensis in the Karoo. South African Journal of Zoology 25: 199–200.
Kerr, P.J. and Donnelly, T.M. 2013. Viral infections of Rabbits. Veterinary Clinics of North America: Exotic Animal Practice. 16 (2): 437–468.
Kryger, U. 2002. Genetic variation among southern African hares (Lepus spec.) as inferred from mitochondrial DNA and microsatellites. Ph.D. Thesis. University of Pretoria.
Kryger, U., Keith, M., Collins, K. and van Jaarsveld, A. 2004. Lepus saxatilis. In: Y. Friedmann and B. Daly (ed.), Red Data Book of the Mammals of South Africa: A Conservation Assessment, pp. 416-417. Endangered Wildlife Trust and CBSGCAMP Programme and Database, South Africa.
Kryger, U., Matthee, C., Keith, M., Collins, K. and van Jaarsveld, A. 2004. Lepus capensis. In: Y. Friedman and B. Daly (eds), Red Data Book of Mammals of South Africa: A Conservation Assessment, pp. 414-415. Endangered Wildlife Trust and CBSG (IUCN/SSC).
Kryger, U., Robinson, T. J. and Bloomer, P. 2004. Population structure and history of southern African scrub hares, Lepus saxatilis. Journal of Zoology (London) 263: 121-133.
Lado, S., Alves, P.C., Islam, M.Z. et al. 2019. The evolutionary history of the Cape hare (Lepus capensis sensu lato): insights for systematics and biogeography. Heredity 123, 634–646.
Maliehe, T.M. 1993. Non-wood forest products in South Africa. Paper prepared for the Regional Expert Consultation on non-wood forest products for English-speaking African countries. Commonwealth Science Council, Arusha, Tanzania.
Melo-Ferreira, J., Boursot, P., Carneiro, M., Esteves, P J., Farelo, L. and. Alves, P.C. 2012. Recurrent Introgression of Mitochondrial DNA Among Hares (Lepus spp.) Revealed by Species-Tree Inference and Coalescent Simulations. Systematic Biology, 61(3): 367.
Ntiamoa-Baidu, Y. 1997. Wildlife and food security in Africa. FAO Conservation Guide 33, Food and Agriculture Organization of the United Nations, Rome. Available at: http://www.fao.org/docrep/w7540e/w7540e00.htm#Contents.
Palacios, F., Angelone, C., Alonso, G. and Reig S. 2008. Morphological evidence of species differentiation within Lepus capensis Linnaeus, 1758 (Leporidae, Lagomorpha) in Cape Province, South Africa. Mammalian Biology-Zeitschrift für Säugetierkunde 73: 358–370.
Petter, F. 1959. Elements d’une revision des lievres africains du sous-genre Lepus. Mammalia 23: 41-67.
Rautenbach, I. L. 1982. Mammals of the Transvaal. Ecoplan monograph, N.S. Supplemento 1: 111-211.
Robinson, T.J., Matthee, C.A. 2005. Phylogeny and evolutionary origins of the Leporidae: a review of cytogenetics, molecular analyses and a supermatrix analysis. Mammal Review 35: 231–247.
Robinson, T.J. 1986. Incisor morphology as an aid in the systematics of the South African Leporidae (Mammalia: Lagomorpha). African Zoology 21: 297–302.
Rocchi, M.S. and Dagleish, M.P. 2018. Diagnosis and prevention of rabbit viral haemorrhagic disease 2. In Practice. 40 (1): 11–16.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Smith, A.T. and Johnston, C.H. 2008. Lepus victoriae.
Smithers, R.H.N. 1971. The mammals of Botswana. National Museums of Rhodesia, Museum Memoir 4: 1-340.
Smithers, R.H.N. and Wilson, V.J. 1979. Check List and Atlas of the Mammals of Zimbabwe Rhodesia. Trustees of the National Museums and Monuments of Rhodesia, Salisbury, Rhodesia.
Stenkewitz, U., Herrmann, E., Kamler, J.F. 2010. Distance sampling for estimating springhare, cape hare and steenbok densities in South Africa. South African Journal of Wildlife Research 40: 87-92.
Stewart, D.R.M. 1971. Diet of Lepus capensis and L. crawshayi. African Journal of Ecology 9: 161–162.
Taylor, P.J. 1998. Regional patterns of small mammal abundance and community composition in protected areas in KwaZulu-Natal. Durban Museum Novitates 23: 42–51.
Suchentrunk, F., Ben Slimen, H. & Kryger, U. 2009. Molecular evidence of conspecificity of South African hares conventionally considered Lepus capensis L., 1758. Mammalian Biology 74(5): 325-343.
Taylor, P.J. 1998. The Smaller Mammals of KwaZulu-Natal. University of Natal Press, Pietermaritzburg, South Africa.
Wessels, T.C. 1978. Bevolkingsdinamika van die vlakhaas Lepus capensis L., 1758 in die Willem Pretorius-wildtuin. M.Sc. Thesis. University of Pretoria.
Wilson, D.E., Lacher, T.E. and Mittermeier, R.A. 2016, Leporidae, pp. 107-148 in Handbook of the Mammals of the World – Volume 6 Lagomorphs and Rodents I, Barcelona :Lynx Edicions on page 131,
World Organisation for Animal Health. Rabbit Haemorrhagic Disease.