

Aardwolf
Proteles cristatus

2025 Red list status
Least Concern
Regional Population Trend
Unknown
Change compared
to 2016
No Change
Overview
Proteles cristatus – (Sparrman, 1783)
ANIMALIA – CHORDATA – MAMMALIA – CARNIVORA – HYAENIDAE – Proteles – cristatus
Common Names: Aardwolf (English), Maanhaarjakkals (Afrikaans), Isambane (Ndebele, Swati, Zulu), Thakadu (Sesotho, Tswana), Xomboni (Tsonga), Thagalu (Venda) Lobo de Tierra (Spanish; Castilian), Protèle (French)
Synonyms: Viverra cristata Sparrman, 1783; Proteles cristatus (Sparrman, 1783) [orth. error]
Taxonomic Note:
Two subspecies are usually recognised: P. c. cristatus from southern Africa, and P. c. septentrionalis from eastern and northeastern Africa. Their validity requires confirmation. Of the recognised two subspecies of Aardwolf, Allio et al. (2021) propose these to be uplifted to species status, i.e. P. cristatus and P. septentrionalis (Allio et al 2021).
Red List Status: LC – Least Concern, (IUCN version 3.1)
Assessment Information
Assessors: Power, RJ.1 & da Silva, J.M.2
Reviewer: Do Linh San, E.3 & Nicholson, S.K.4
Contributor: Patel, T.4
Institutions: 1North West Provincial Government Directorate of Biodiversity Management, 2South African National Biodiversity Institute, 3Sol Plaatjie University, 4Endangered Wildlife Trust
Previous Assessors and Reviewers: de Vries, L., Marneweck, D., Dalerum, F., Page-Nicholson, S., Mills, M.G.L., Yarnell, R., Sliwa, A. & Do Linh San, E.
Assessment Rationale
This species is listed as Least Concern as it is widespread within the assessment region and present in numerous protected areas and habitats, as long as there are termites available. Aardwolves can occupy open and degraded grassland where there is high termite abundance. The expansion of wildlife ranching may be increasing suitable habitat across the assessment region, although this should be weighed against possible increases in direct or indirect persecution. Additionally, climate change is predicted to potentially decrease food availability for the Aardwolf, and it is possible that it will become threatened in the near future. For now, there is no evidence to indicate a range-wide decline.
Regional population effects: There is a continuous range that extends outside of the assessment region into neighbouring countries, and the assessment region does not appear to be a sink. Aardwolves are believed to be good dispersers as they walk tirelessly and can easily cover 20 km in one night when advertising mating rights (Sliwa 1996), and when translocated they tend to travel far, i.e. 35 km (Bothma 1971).
Reasons for Change
Reason(s) for Change in Red List Category from the Previous Assessment: No change
Red List Index
Red List Index: No change
Recommended Citation: Power RJ & da Silva JM. 2025. A conservation assessment of Proteles cristatus. In Patel T, Smith C, Roxburgh L, da Silva JM & Raimondo D, editors. The Red List of Mammals of South Africa, Eswatini and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Regional Distribution and occurrence
Geographic Range
The Aardwolf has a disjunct distribution in Africa, occurring in two discrete areas (1,500 km apart), one in East and northeastern Africa and one in southern Africa (Coe & Skinner 1993; Skinner & Chimimba 2005). The southern African distribution includes South Africa, Eswatini, Namibia, Botswana, Zimbabwe and a narrow strip in the western region of Mozambique. Aardwolf distribution is largely determined by the distribution of Trinervitermes termites, which constitute their main food (Anderson 2013).
Within the assessment region, Aardwolves occur throughout the nine provinces in areas with suitable habitats (Richmond-Coggan 2014; De Vries et al. 2016). Richmond-Coggan (2014) interviewed 190 landowners across South Africa and asked whether they had seen Aardwolf on their properties. Aardwolves were found on at least one property in all nine of South Africa’s provinces. Forty-five percent of respondents indicated that this species was present on their land. Eastern Cape (50%), Free State (71%), Gauteng (50%), and Northern Cape (50%) respondents indicated the highest proportion of properties with Aardwolf presence. Lowest Aardwolf presence was indicated in KwaZulu-Natal (25%). This species favours drier areas, most notably in the Northern Cape, where it has always occurred in the Northern Cape (Stuart 1981; Lloyd & Millar 1984). They occur in the Tswalu Kalahari Reserve (Smith et al. 2023).
The Aardwolf occurs across the whole of the North West Province (Power et al. 2019), and are most common in the southern and western parts (Power 2014). In the Ngaka Modiri Molema District, around Mafikeng, just over 30% of Game Farmers had reported Aardwolf based on questionnaires (NWPG 2012), while most had reported them from the far west of the province (NWPG 2013). In the Northern Bojanala Platinum District of the North West Province, the Aardwolf occurred in less than half the vegetation types across this Bushveld district (NWPG 2011). Aardwolf are present in the Mankwe Wildlife Reserve east of Pilanesberg (Hart et all 2022), Madikwe (Smith et al. 2023), and they have been reported of late as roadkill occurrence records across the known areas of occurrence (NWPG 2024).
In the Western Cape, the Aardwolf was known from most of the Karoo parts of the Western Cape (Von Richter 1972; Stuart 1981; Lloyd and Millar 1984; Woodgate et al. 2023), and they still occur across parts of the Karoo (Schurch et al. 2021), as well as locally in the Fynbos areas too, albeit in lower abundances (Nieman et al. 2021). In the Boland Mountain Complex, Aardwolf are reported from 4% of properties (Nieman et al. 2021) and are locally rare, as it is largely suboptimal habitat there. Aardwolf have always occurred over much of the Eastern Cape, especially the inland Karoo areas (Von Richter 1972; Stuart 1981; Lloyd and Millar 1984). They occur in the Mountain Zebra National Park (Smith et al. 2023). Aardwolf have always been common and widespread in the Free State (Von Richter 1972; De Vries et al. 2016), and they still seem present throughout.
The Aardwolf is known from most of the western half of KwaZulu-Natal (Rowe-Rowe 1992). By the 1970s there had been a few sight records on the margins of Maputaland in northern Kwazulu-Natal of the Aardwolf (Pringle 1977). They appeared absent from most of Zululand, and Maputaland, given several intensive carnivore-focussed camera trap surveys, which did not detect the species (see Ramesh et al. 2017; Da Rosa 2019; Pretorius 2019). The species may have retracted from the coastal plain of Zululand. The Aardwolf has always been rare in the Kruger National Park and Lowveld as a whole, but readily along the escarpment (Von Richter 1972), and occur on the Highveld there, i.e. Telperion Nature Reserve (Webster et al. 2021). Further east, they occur also in the Verloren Vallei Nature Reserve (Loock & Swanepoel 2019). The Aardwolf occurs throughout the Bushveld of Limpopo, but less so in the Lowveld (Von Richter 1972; Skinner & Chimimba 2005).
Aardwolf have been recorded in Eswatini, particularly the western Highveld part (Monadjem 1998). There are no records from Lesotho (Lynch 1994).
The distribution is similar to the last assessment, with perhaps a reduction from north-eastern KwaZulu-Natal (see Map).
Elevation / Depth / Depth Zones
Elevation Lower Limit (in metres above sea level): 0
Elevation Upper Limit (in metres above sea level): 2000
Depth Lower Limit (in metres below sea level): (Not specified)
Depth Upper Limit (in metres below sea level): (Not specified)
Depth Zone: (Not specified)
Map
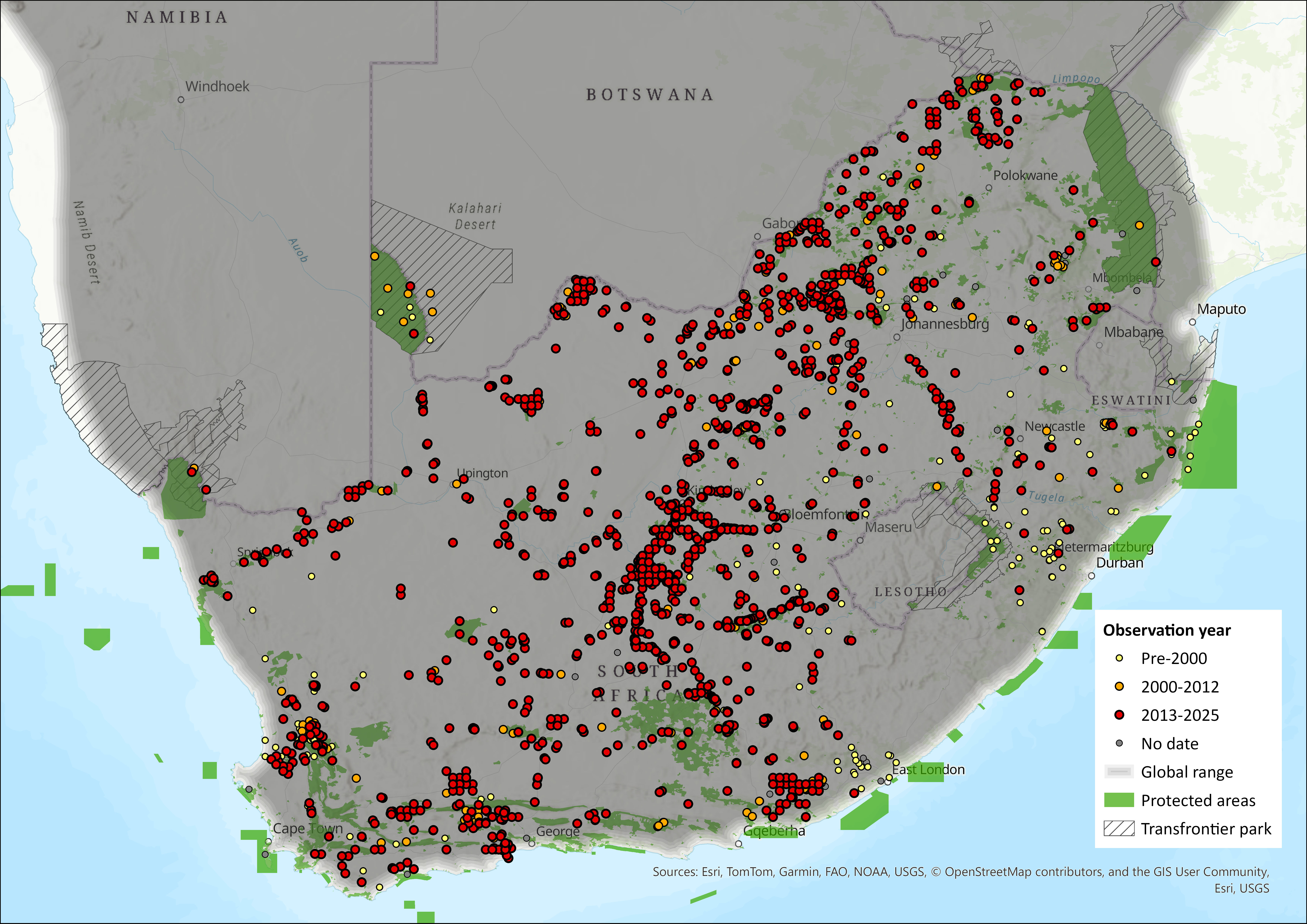
Figure 1. Distribution records for Aardwolf (Proteles cristatus) within the assessment region (South Africa, Eswatini and Lesotho). Note that distribution data is obtained from multiple sources and records have not all been individually verified.
Biogeographic Realms
Biogeographic Realm: Afrotropical
Occurrence
Countries of Occurrence
| Country | Presence | Origin | Formerly Bred | Seasonality |
| Angola | Extant | Native | – | – |
| Botswana | Extant | Native | – | – |
| Djibouti | Presence Uncertain | Native | – | – |
| Egypt | Extant | Native | – | – |
| Eritrea | Extant | Native | – | – |
| Eswatini | Extant | Native | – | – |
| Ethiopia | Extant | Native | – | – |
| Kenya | Extant | Native | – | – |
| Lesotho | Presence Uncertain | Native | – | – |
| Mozambique | Extant | Native | – | – |
| Namibia | Extant | Native | – | – |
| Somalia | Extant | Native | – | – |
| South Africa | Extant | Native | – | – |
| Sudan | Extant | Native | – | – |
| Tanzania, United Republic of | Extant | Native | – | – |
| Uganda | Extant | Native | – | – |
| Zambia | Extant | Native | – | – |
| Zimbabwe | Extant | Native | – | – |
Large Marine Ecosystems (LME) Occurrence
Large Marine Ecosystems: (Not specified)
FAO Area Occurrence
FAO Marine Areas: (Not specified)
Climate change
It is important to consider a species’ ecological and biological traits to determine the likely mechanisms of climate change impact and quantify this using expert knowledge (Foden & Young 2016). An increase in shrub cover as predicted by many climate change models (Foden and Young 2016), would negatively impact the Aardwolf, given it is a species of open areas and would benefit from any situation where drying out takes place.
As a highly specialised forager, Aardwolves may be vulnerable to future environmental changes, for example, shifts in grassland communities caused by global warming. However, as the ecological outcomes of future temperature fluctuations are uncertain, it is unclear to what extent they will impact Aardwolf populations (De Vries et al. 2016).
Population information
In prime habitat (open grassland and scrub regions), densities may reach 1 adult / km² on farms with good populations of termites and no persecution by farmers (Bashant 2008; Anderson 2013).
In conducting a feasibility assessment aimed at the Cape Floristic Region in southern South Africa, based on literature, an Aardwolf density estimate for the area was 11.1 Aardwolves / 100 km² (Boshoff et al. 2002). Similarly, shortly after, a total population size of 11,716 was estimated for the species there (Kerley et al. 2003). This was however indirectly done on habitat suitability, and they were not directly enumerated.
In the North West, Aardwolf are most common in the western Kalahari and southern grasslands on the Highveld than anywhere else in the province. The species was very common at Bloemhof Dam Nature Reserve (Power 2014).
In surveys assessing the impact of hunting and disturbance on Aardwolf inter alia, the control site (!Khamab Kalahari Reserve) in 2015, revealed a photo-capture rate (Relative Abundance Indices – RAI) of 0.6±1.1 foxes/100 trap nights, and none were recorded on adjacent hunting farms adjacent the reserve (NWPG 2016). A year later, at another benchmark reserve (Molopo), the RAI was higher at 5.7±12.9/100 trap nights, and naïve occupancy was 22% there, which is in tandem with an increasing population of 14% (NWPG 2017). An ecologically comparable livestock farming area registered an RAI of 0.8±1.7 (naïve occupancy of 17%), while none were recorded on a hunting ranch (NWPG 2017). Although RAIs have its shortfalls, these findings do suggest at least in this area protected areas have higher abundances of this species.
However, in the Western Cape Karoo, the opposite was the case there where their relative abundance was higher for livestock farming systems when compared to wildlife-friendly farms, or protected areas (Schurch et al. 2021; Woodgate et al. 2023). In Nuweveld mountains of the Western Cape Karoo, with a change of management to wildlife-friendly farming, Aardwolf steadily decreased in abundance up until 2019 over a four-year period (Schurch et al. 2021).
Between 2013 and 2015, camera trapping took place in the Western Cape Karoo and the Aardwolf registered a higher occupancy on commercial small livestock farmlands, than ecologically comparable private and state protected areas (Woodgate et al. 2023). Similarly, then for these nearby Karoo ecosystems, the livestock farming properties offered better prospects for the Aardwolf, likely due to higher stocking rates and slightly degraded rangelands.
Elsewhere a high number of detections was found in the Mountain Zebra National Park (Smith et al. 2023), while low number of detections were reported for Madikwe Game Reserve (Smith et al. 2023), Tswalu Kalahari Reserve (Smith et al. 2023), and the Verloren Vallei Nature Reserve (Loock & Swanepoel 2019). Although these are detections on camera traps and can only enable an appraisal of relative abundance.
Our understanding of the population sizes and trends of Aardwolves is uncertain and further research is required to quantify the population.
Population Information
Current population trend: Unknown
Continuing decline in mature individuals? Unknown
All individuals in one subpopulation: Unknown
Number of mature individuals in population: Unknown
Number of mature individuals in largest subpopulation: Unknown
Number of Subpopulations: Unknown
Extreme fluctuations in the number of subpopulations: Unknown
Continuing decline in number of subpopulations: Unknown
Severely fragmented: No. Although this species seems to be largely dependent on the presence of harvester termites, its distribution is relatively continuous as favourable habitats are generally connected. In addition, the Aardwolf has the ability to cover large distances (Sliwa 1996) and thus connect between fragments – if they occur.
Quantitative Analysis
Probability of extinction in the wild within 3 generations or 10 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 5 generations or 20 years, whichever is longer, maximum 100 years: (Not specified)
Probability of extinction in the wild within 100 years: (Not specified)
Population genetics
Westbury et al. (2019) presented the first complete Aardwolf mitochondrial genome and provided novel information about their evolutionary origins and genetic structure within a single population. Through the investigation of complete mitochondrial genomes from five individuals from a single population within South Africa, they found the mitochondrial diversity of this population to be moderate when compared to a number of other mammals (Westbury et al. 2019).
High levels of genetic diversity and more stable population sizes through time are seen in the Aardwolf (Westbury et al. 2021).
Allio et al. (2021) used roadkill samples to study the genomic diversity of the Aardwolf, specifically of the disjunct distributions in Eastern and Southern Africa (Allio et al. 2021).
By comparing their genome-wide genetic differentiation, the two subspecies of Aardwolf might warrant species status (P. cristatus and P. septentrionalis) (Allio et al 2021). Information on genetic structure and heterozygosity in the Aardwolf has been determined, including historical effective population sizes inter alia (Allio et al. 2021). Contemporary Ne has not been quantified for the species, however based on estimates provided by Kerley et al. (2003; i.e., 11,716), it can be inferred using a conversion ratio between 0.1-0.3. This equates to at least 1,172-3,515, exceeding the threshold of 500 which suggests a healthy, stable population. These values should be verified using population genetic techniques.
Habitats and ecology
Open, grassy plains constitute the prime habitat of this species. The Aardwolf is entirely absent from forests or pure desert (Stuart 1981; Anderson 2013). However, in southern Africa it occupies diverse habitats, ranging from the karroid habitats of the Western Cape and Eastern Cape (Lloyd & Millar 1984; Stuart 1981), the grasslands and scrub of Botswana, the open savannah woodlands of Zimbabwe, to the inland gravel plains of the Namib Desert in Namibia (Skinner & Chimimba 2005). The species has been recorded to 2,000 m asl in Ethiopia (Yalden et al. 1996). The Aardwolf is an inhabitant of open country, even lowland Fynbos, and in Namaqualand uses broken country too (Stuart 1981). The Aardwolf avoids forest and valley bushveld (Stuart 1981). Aardwolves were recorded in open grassland areas supporting numerous Trinervitermes mounds (Matsebula et al. 2009; Power 2014), and even in mountain grasslands (Monadjem 1998) and degraded grassland (Power 2014).
Throughout their distribution, Aardwolves have been recorded to feed primarily on Nasute Harvester Termites (genus Trinervitermes) (Kruuk and Sands 1972; Cooper and Skinner 1979; Richardson 1987a; Anderson 1994; Matsebula et al. 2019), and, in any particular region, mainly on one species, while there may be trace amounts of other genera of termites, ants, other insects, millipedes (Cooper and Skinner 1979) and even sun spiders and scorpions (De Vries 2011). No vertebrate remains or traces of carrion were found in the diet of Aardwolf (Cooper and Skinner 1979). The dentition and jaw musculature of the Aardwolf is said to be incapable of taking livestock (Stuart 1981; Skinner & Chimimba 2005). However, the Aardwolf is capable of exploiting alternative food sources and occasionally do so (Yarnell and Mactavish 2013). Unlike the other typical “anteaters”, Aardwolves do not dig for their prey but lick foraging termites off the soil surface. Consequently, they do not possess the physical features such as a digging apparatus which are characteristic of typical ant- and termite-eating mammals (Anderson 1994). The nutritional quality of T. trinervoides, like other invertebrates, is poor. They compensate for this by consuming vast quantities of termites. Aardwolves consumed between 222, 445 Trinervitermes (Williams et al. 1997) to up to 300,000 termites per night during the warm summer months (Anderson 1994). It is predicted that Aardwolves consume termites at an intake rate of 33 termites per second while feeding (Williams et al. 1997). During winter, they consumed 5,867 Hodotermes and 2,515 Trinervitermes per night (Williams et al. 1997).
Survival during winter, however, is achieved through a combination of behavioural and physiological adaptations. As a consequence of the Aardwolf’s specialisation on this unique diet, and the ability to cope with the absence of termites during winter, it has invaded a trophic niche which is almost entirely devoid of competitors. The body masses of Aardwolves decrease significantly seasonally (Anderson 1994; Van Jaarsveld et al. 1995), by 25% from approximately 10 kg to 7,5 kg during the winter months (April – August) (Anderson 1994), and they are at their lowest at the end of winter (August) (Van Jaarsveld et al. 1995). This is a nutritionally stressful period for Aardwolves, as evidenced by increased juvenile mortality, a 20% decline in body mass and depletion of subcutaneous fat reserves (Anderson 1980). The Aardwolf’s metabolic rate is 11% lower during the winter months, resulting in reduced energy requirements during the period of limited food availability (Anderson 1980).
The species may however be an opportunist, and the observed behavioural plasticity could be advantageous under specific environmental conditions (De Vries 2011; Yarnell and Mactavish 2013). There are sometimes significant seasonal fluctuations in the diet of Aardwolves (Kruuk & Sands 1972; Richardson 1987b; Taylor & Skinner 2000). In Eswatini, Hodotermes contributed very little to the diet of Aardwolves there and were only recorded in the wet season (Matsebula et al. 2009).
Compared to other myrmecophagous mammals, Aardwolf utilised the fewest prey categories and had the lowest dietary niche breadth (Pietersen and Robertson 2023), reinforcing how specialised they are. The Aardwolf is therefore well adapted to feed on a diet of Trinervitermes termites, a food niche that has been largely unexploited by other animals, and it uses various ecological, behavioural and physiological adaptations to cope with this seasonally unavailable food source (Anderson 1980).
All scientific reports on Aardwolf foraging and diet agree that this species is an obligate insectivore (Cooper & Skinner 1979; Richardson 1987a; Matsebula et al. 2009; de Vries et al. 2011) and poses no risk to livestock. There are, however, historic anecdotal accounts of Aardwolf taking vertebrate prey items (Estes 1991; Nowak 1991; Yarnell & Mactavish 2023). For example, Kingdon (1977) reported that an Aardwolf had killed a number of birds and Bothma and Nel (1980) described Aardwolves to have eaten small rodents, carrion, eggs, birds and baby tortoises. Yarnell and Mactavish (2013) describe how a captive goose was killed by an Aardwolf. However, such instances are likely to be very rare based on the paucity of such reports (Yarnell & MacTavish 2013).
They are largely independent of water, obtaining their moisture requirements from termites (Anderson 2013).
Female Aardwolves come into pro-oestrus in the latter weeks of June and mating generally occurs in the first two weeks of July (Skinner & Chimimba 2005). Aardwolves are considered socially monogamous but sexually polygamous with numerous extra-pair copulations occurring in the short winter mating period (Richardson 1987c; Kotze et al. 2012; Marneweck et al. 2015; Skinner & Chimimba 2005). Sometimes females practice regular promiscuous matings, as a result of which some males would appear to be cuckolding their neighbours (Richardson 1987b; Kotze et al. 2012). However, mate desertion by males may also occur in response to female promiscuity (Richardson and Coetzee 1988). A cuckolded male Aardwolf may still be raising some of his own offspring and should desert only on the rare occasions when an alternative receptive female is exclusively available (Richardson and Coetzee 1988).
Gestation period is approximately 90 days, and most litters consist of between two to four cubs, sometimes five. Aardwolves have a relatively short gestation period and endure extensive postnatal investment in their cubs, perhaps in response to potentially excessive juvenile mortality (Van Jaarsveld 1993). In the assessment region, cubs are born between October and December (Skinner & Chimimba 2005; Marneweck et al. 2015). The female gives birth in a den where the cubs remain for up to a month before emerging. Males will assist in the rearing of the young. Between 4 and 6 weeks, the cubs remain close to the den. As the cubs get older, they will venture further away from the den. Eventually between 12 and 16 weeks, the cubs will forage with an adult throughout the territory (Skinner & Chimimba 2005). Females have an extended lactation period and there is intensive parental care (Van Jaarsveld et al. 1995).
Aardwolves are nocturnal and spend the daylight hours underground (Richardson and Anderson 2005). When inactive, Aardwolves sought refuge in underground dens, which were usually modified springhare Pedetes capensis burrows. Daily denning times during winter were approximately 20 hours in comparison to a shorter 15 hours 20 min during summer (Anderson 1994). While inactive, the Aardwolf lowers its body temperature to an average of 34.1°C during the middle of the inactive period during winter and in some individuals, this results in energy savings of up to 18% (Anderson 1980). In the Aardwolf, evaporative water loss rates are low, and are further reduced during the winter months, typically adaptations to survive in a desert environment and on a seasonally unavailable food source (Anderson 1980).
Aardwolves undergo seasonal changes in pelage density and thermal conductance allow for more passive heat transfer during summer and improved heat retention during winter (Anderson 1980).
In Aardwolf, evaporative water loss increased with air temperature for both seasons but was higher in summer than winter (Anderson et al. 1997). A time budget for active periods showed that Aardwolves spent a significantly larger proportion of their active time feeding in summer than in winter (26.7 vs. 15.2%) (Williams et al. 1997).
Aardwolves were active for an average of 8.77 hours per 24-hour period in summer, but only 4.05 hours during winter (Williams et al. 1997). Aardwolves seem to forage for termites earlier in the night than sympatric Bat-eared Foxes (Otocyon megalotis) during the colder months to lower the risk of losing access to termite colonies (Smith et al. 2023).A time budget for active periods showed that Aardwolves spent a significantly larger proportion of their active time feeding in summer than in winter (26.7 vs. 15.2%) (Williams et al. 1997).
During winter, Aardwolves spent 12.6% of their time engaged in in reproductive activities, i.e. courtship and copulation (Williams et al. 1997).
Scent marking by the Aardwolf help to maintain and ensure, territorial integrity of a resident’s home-range (Richardson 1991) and are unlikely to entirely exclude intruders from exploiting resources within a territory, they may however limit the degree of time and extent that a territory is intruded into and hence indirectly protect its resources (Richardson 1993). Aardwolves scent marked from anal glands to mark their territories (Sliwa 1996), and scent marking may function in intimidating intruders and to synchronize mating (Sliwa and Richardson 1998). By defending an exceptionally large territory, as well as having two females, weakened one male Aardwolf’s ability to guard his female partners efficiently from Extra Pair copulations (Sliwa 1996).
A comprehensive review of the species’ ecology can be found in Koehler and Richardson (1990) and Anderson (2013).
Ecosystem and cultural services: Since the Aardwolf is dietary specialist of termites, mostly Trinervitermes (Kruuk and Sands 1972; Cooper and Skinner 1979; Anderson 1994; Matsebula et al. 2019), it can surely be of benefit to farmers who utilise rangelands, given use of a prey item which competes with their own livestock for grazing.
IUCN Habitats Classification Scheme
| Habitat | Season | Suitability | Major Importance? |
| 2.1. Savanna -> Savanna – Dry | – | Suitable | – |
| 2.2. Savanna -> Savanna – Moist | – | Suitable | – |
| 3.5. Shrubland -> Shrubland – Subtropical/Tropical Dry | – | Suitable | – |
| 3.6. Shrubland -> Shrubland – Subtropical/Tropical Moist | – | Suitable | – |
| 3.8. Shrubland -> Shrubland – Mediterranean-type Shrubby Vegetation | – | Marginal | – |
| 4.4. Grassland -> Grassland – Temperate | – | Marginal | – |
| 4.5. Grassland -> Grassland – Subtropical/Tropical Dry | – | Suitable | – |
Life History
Generation Length: (Not specified)
Age at Maturity: Female or unspecified: (Not specified)
Age at Maturity: Male: (Not specified)
Size at Maturity (in cms): Female: (Not specified)
Size at Maturity (in cms): Male: (Not specified)
Longevity: (Not specified)
Average Reproductive Age: (Not specified)
Maximum Size (in cms): (Not specified)
Size at Birth (in cms): (Not specified)
Gestation Time: (Not specified)
Reproductive Periodicity: (Not specified)
Average Annual Fecundity or Litter Size: (Not specified)
Natural Mortality: (Not specified)
Does the species lay eggs? (Not specified)
Does the species give birth to live young: (Not specified)
Does the species exhibit parthenogenesis: (Not specified)
Does the species have a free-living larval stage? (Not specified)
Does the species require water for breeding? (Not specified)
Movement Patterns
Movement Patterns: (Not specified)
Congregatory: (Not specified)
Systems
System: Terrestrial
General Use and Trade Information
There are documented accounts of Aardwolves being consumed as food or used in medicinal practices by indigenous tribes in Africa (Richardson 1984; Koehler & Richardson 1990; Hofer & Mills 1998). It is likely that such uses and practices are limited in the assessment region (De Vries et al. 2016)
Body parts of the Aardwolf are traded in the Eastern Cape (Simelane & Kerley 1998), Gauteng (Whiting, Williams & Hibbits 2016) and in the Western Cape (Nieman et al. 2018). The species appeared in 5% of herbalist shops in the Eastern Cape (Simelane & Kerley 1998).
In the Western Cape at least, the entire skin of the Aardwolf can be used decoratively and as clothing (Nieman et al. 2018) and can sell for R2000-R6000 (Nieman 2018). A skin piece and the bones of the Aardwolf can be used to attract good fortune (Nieman et al. 2018) and can be sold for R1000 – R1700 (Nieman 2018).
Hunting: The species is trophy hunted but it does not seem to be significant, however, an increase is evident (Professional Hunters register from 2010 to 2019). The most important provinces hunting the species are KwaZulu-Natal, followed by the Eastern Cape and Northern Cape. In the North West, since 2020 there have been 5 issued permits to hunt the species (NW Directorate of Biodiversity Management records, 2020-2023).
Wildlife ranching is generally thought to preserve and potentially expand suitable habitat for this species (but see Schurch et al. 2021; Woodgate et al. 2023), since Aardwolves thrive on open and moderately degraded grassland with high termite abundance (for example, Power 2014). Landowner attitudes are also generally positive towards this species, although it is sometimes mistakenly persecuted as a damage-causing animal (De Vries et al. 2016).
Ecotourism: The species is endearing to see, and most members of the public can be afforded the best viewing by embarking on a guided night drive to see them with a spotlight. Good Parks to see them are the Mokala National Park, Karoo National Park and Mountain Zebra National Park. Several private reserves in the Northern Cape have risen to the challenge and offer night viewing safaris to see the species, such as Marrick Safari Lodge, and the KhoiSan Karoo Conservancy.
| Subsistence: | Rationale: | Local Commercial: | Further detail including information on economic value if available: |
| Yes | – | – | – |
National Commercial Value: No
International Commercial Value: No
| End Use | Subsistence | National | International | Other (please specify) |
| 1. Food – human | true | – | – | – |
| 3. Medicine – human & veterinary | true | – | – | – |
Is there harvest from captive/cultivated sources of this species? (Not specified)
Harvest Trend Comments: (Not specified)
Threats
Persecution as a problem animal
In South Africa, the Aardwolf was previously persecuted by some farmers for the mistaken belief that it was a predator of livestock, chickens and eggs (Hey 1964; Von Richter 1972; Richardson 1984; Anderson 1988). However, such reports are not substantiated by studies of gut or faecal contents and probably result from mistaken identity with hyaenas or jackals (Anderson 2013), and this perception has subsequently changed. They are, however, the occasional inadvertent victims of problem animal control operations, especially those using gin traps (M.D. Anderson pers. comm. 2015, In: De Vries et al. 2016).
Aardwolves have been reported to have killed a captive goose at (Yarnell and MacTavish 2013). Aardwolves are common on farmland and are likely to encounter farms and domestic poultry frequently, suggesting that if poultry predation was typical, one would expect many more records to exist (Yarnell and MacTavish 2013). In the North West Province, DCA officers respond to one complaint for the Aardwolf once per annum at most (North West Biodiversity Management Directorate records). In one case, and emaciated juvenile was obtained near Mafikeng, and taken to a rehabilitation centre, but it succumbed, and died.
Since the last assessment, there was one report of an illegally killed Aardwolf (NorthWest Biodiversity Management Directorate records, 2023). Although these two deaths can be reported upon, illegal killing probably happens more often than this.
Dogs
Of three Aardwolves monitored in the North West by conservation authorities during the 1960s’ two of them were killed by dogs (Bothma 1971).
Roads
There have been several roadkill records from the Northern Cape (Bullock et al. 2011) and recently in the North West (NWPG 2024), and elsewhere (Allio et al. 2021). Several of the distribution records which are linearly arranged across central South Africa glaringly point to this being a threat at least. In northern Limpopo, Collinson et al. (2015) drove a total of 14,400 km over 120 days spanning 10 months across 2011 and 2012, on a 120 km circuit around the Venetia Limpopo Nature Reserve, and a stretch that abutted the Mapungubwe National Park. On this circuit, two Aardwolf were reported to have been killed on these roads (Collinson et al. 2015). Given the 10-month study period, this would amount to 2.4 Aardwolf killed by roads per annum there.
More recently, in the North West, on a commissioned study, of 81,673 km driven on randomly driven roads of the North West Province from 2020 to 2024, there were 5 Aardwolf roadkill recorded (NWPG 2024, and previous annual reports from 2020). Locally, in the same province, on a systematic Lichtenburg-Mahikeng-Ottoshoop road kill survey (150 km circuit), conducted from 2021 to 2024, 13,200 km was driven, and the circuit was driven on 100 occasions, there were no mortalities recorded of the Aardwolf over this time period (NWPG 2024, and previous annual reports), despite the species occurring in the area.
These reports however do not indicate roadkill to be excessive, though it should not be overlooked.
Body parts uses and trade
The body parts and medicinal uses trade has increased in the Western Cape, though compared to other species, it was not significantly used, and was not subject to an assessment of this as a threat (Nieman et al. 2018).
Habitat degradation and loss
Loss of habitat, through urbanisation or expansion of industrial agriculture may have negative impacts. Additionally, some farmers in South Africa destroy termitaria, using a plough or poisons, which diminishes Aardwolves’ resource base, and poisons used for locust control may also have negative effects (Anderson 2013). Based on habitat transformation in the Cape Floristic Region, the Aardwolf population was said to have declined by 30.6 % (Kerley et al. 2003).
Disease
Rabies in wildlife species followed different spatial distributions where Aardwolf cases were more frequent in southern and western South Africa (Koeppel et al. 2022).
Pesticides
Pesticides that aim to poison their food source can have an impact on them (Ray et al. 2005).
Global threats
A global threat category assesses how each species is differentially impacted by a set of ten external threats commonly encountered by African carnivores (see Ray et al. 2005). Each species was subjected to a scoring process for each threat that assessed the relative severity (its effect on the species), urgency (timescale over which it is most likely to occur), probability (likelihood of occurrence), and geographical extent of the threat through the species’ range in Africa (modified from Coppolillo et al. 2004, In: Ray et al. 2005). The Aardwolf scored 46, and insect control was one the threats it faces in its global range (Ray et al. 2005), and this was on s a scale relatively moderate.
Conservation
Aardwolves are found in numerous well-managed protected areas across their distribution range, including the following that are important owing to high densities there, and stable populations: Tswalu Kalahari Reserve, Mokala National Park (Northern Cape Province), Mountain Zebra National Park, Greater Addo Elephant National Park, Great Fish River Nature Reserve (Eastern Cape Province), Karoo National Park (Western Cape), Bloemhof dam Nature Reserve, SA Lombard Nature Reserve, Botsalano Nature Reserve (North West), Tussen-die-Riviere Nature Reserve (Free State). Therefore, it seems that medium-sized protected areas (< 100, 000 ha) are important in conserving this species.
Although they occur in Kruger National Park (Mpumalanga and Limpopo provinces), Pilanesberg National Park (North West Province), Kgalagadi Transfrontier Park (De Vries et al. 2016), they are rare there in these protected areas, perhaps due to high densities of predators.
Grassland burning and livestock grazing may increase populations of Trinervitermes, which would therefore benefit Aardwolves (Kruuk and Sands 1972; Anderson 2013).
In the North West Province of 609 Game Farms assessed, 30% of them had exceeded an ecological carrying capacity (Power 2014). This degraded habitat would likely be conducive to Aardwolf occupation.
Educational campaigns could be put in place to dispel the myth that Aardwolves prey on livestock. In addition, landowners must be able to identify Aardwolves, the areas which they favour and know the general biology of the species, particularly what they eat. Game and nature reserves also need to be incorporated into such campaigns, as some managers know very little about Aardwolves. Farmers tend to confuse this species with the Brown Hyaena (Parahyaena brunnea) largely owing to its Afrikaans name of Maanhaarjakkals, which renders it similar in appearance to the Brown Hyaena. Educational material highlighting the differences in a popular magazine should quell this misunderstanding (Power 2014), while they also do with jackals too. The De Larey Branch of the South African Hunters and Game Conservation Association are to embark on a training exercise to encourage the use of spotters to identify target animals when on jackal control operations, given the Aardwolf is mistakenly shot on occasion. This is to reduce accidental killing of non-target ecologically useful, and cryptic species.
Injured Aardwolves treated in rehabilitation have been released and monitored, though none of them did survive, one attempted to home back to its origin. Three translocated Aardwolves that were monitored in the North West travelled an average of 16.1 km, with a maximum distance of 35.km travelled in 24 days (Bothma 1971).
Recommendations for land managers and practitioners:
- Burn grasslands to maintain termite density.
- Maintain termite density by not destroying termitaria, using a plough or poisons.
- Maintain grazing regimes in open grassland.
- Design and maintain a specific Aardwolf education webpage – so far there is too little information centrally available.
Research priorities:
- Conduct genetic studies to evaluate large-scale population structure and movement across the species’ range.
- Conduct genetic research into subspecies validity or even presence of distinct, but so far unrecognised, subspecies or populations.
- Quantify population trends and sizes. Despite its pervasiveness in basic research on hyenas and aardwolves, individual identification has rarely been applied to the management of these species. Individual identification using naturally occurring markings in applied research could prove immensely helpful, as this could further improve accuracy of density estimates, reveal characteristics of suitable habitat, identify threats to population persistence, and help to identify individual problem animals (Spagnuolo et al. 2022).
- Evaluate the ecological impacts of Aardwolves in grassland communities, both as predators and prey.
- Determine suitable burning regimes which enhance termite abundance. Despite the ecological importance of the Hyaenidae, and the threats they face globally, there has been no synthesis of management-related literature regarding these species (Wilkinson et al. 2024).
- Conduct a detailed and extensive survey specifically designed for Aardwolves in order to get a better representation of the species’ finer-scale distribution.
- The most- studied broad topics regarding aardwolves were ecology/diet and endocrinology/morphological development/anatomy, while the least- studied broad topics were disease, interspecific interactions and human impacts/ conflicts/benefits (Wilkinson et al. 2024).
Encouraged citizen actions:
- Report sightings on virtual museum platforms (for example, iNaturalist and MammalMAP), especially outside protected areas.
- Encourage landowners and ranchers to sustain termites on their farms by maintaining open grassland communities.
- Educate farmers on Aardwolf feeding ecology.
Bibliography
Allio R, Tilak MK, Scornavacca C, Avenant NL, Kitchener AC, Corre E, Nabholz B, Delsuc F. 2021. High-quality carnivoran genomes from roadkill samples enable comparative species delineation in aardwolf and bat-eared fox. E-life 18;10:e63167.
Anderson MD. 1980. Aardwolf adaptations: A review: Animals. Transactions of the Royal Society of South Africa 59(2):99-104.
Anderson, M.D. 1988. The aardwolf – harmless “ant-eater”. Farmers Weekly August 19: 34-36.
Anderson MD. 1994. The influence of seasonality and quality of diet on the metabolism of the aardwolf, Proteles cristatus (Sparrman 1783). University of Pretoria, Pretoria.
Anderson, M.D. 2013. Proteles cristatus. In: J. Kingdon and M. Hoffmann (eds), The Mammals of Africa. Volume V: Carnivores, Pangolins, Equids, Rhinoceroses, pp. 560. Bloomsbury Publishing, London.
Bashant J. 2008. An assessment of spotlight distance sampling for estimating density of nocturnal carnivores. B.Sc. Honours Thesis. University of Pretoria.
Boshoff AF, Kerley GI, Cowling RM. 2002. Estimated spatial requirements of the medium-to large-sized mammals, according to broad habitat units, in the Cape Floristic Region, South Africa. African Journal of Range and Forage Science. 19(1):29-44.
Bothma JD. 1971. Notes on the movement of black-backed jackal and the aardwolf in the western Transvaal. African Zoology.;6(2):205-207.
Bothma J du P, Nel JAJ. 1980. Winter food and foraging behaviour of the aardwolf Proteles cristatus in the Namib–Naukluft Park. Madoqua 12: 141–145.
Bullock KL, Malan G, Pretorius MD. 2011. Mammal and bird road mortalities on the Upington to Twee Rivieren main road in the southern Kalahari, South Africa. African Zoology 46(1):60-71.
Coe MJ, Skinner JD. 1993. Connections, disjunctions and endemism in the eastern and southern African mammal faunas. Transactions of the Royal Society of South Africa. 48(2):233-55.
Collinson WJ, Reilly BK, Parker DM, Bernard RT, Davies-Mostert HT. 2015. An inventory of vertebrate roadkill in the greater Mapungubwe Transfrontier conservation area, South Africa. African Journal of Wildlife Research 45(3): 301-311.
Cooper RL, Skinner JD. 1979. Importance of termites in the diet of the aardwolf Proteles cristatus in South Africa. South African Journal of Zoology 14: 5–8.
Da Rosa BRP. 2019. Optimizing Small Mammal Relative Abundance Measures Using Non-Invasive Sampling and Assessment of its Contribution to Occupancy Modelling of Small Carnivores in Dry Woodland Savannah of South Africa. Master’s thesis, University of Lisbon, Lisbon.
De Vries L. 2011. Impacts of a specialist diet on aardwolf ecology. Msc dissertation, University of Pretoria, Pretoria.
De Vries JL, Marneweck D, Dalerum F, Page-Nicholson S, Mills MGL, Yarnell RW, Sliwa A, Do Linh San E. 2016. A conservation assessment of Proteles cristata. In Child MF, Roxburgh L, Do Linh San E, Raimondo D, Davies-Mostert HT, editors. The Red List of Mammals of South Africa, Swaziland and Lesotho. South African National Biodiversity Institute and Endangered Wildlife Trust, South Africa.
Foden WB, Young BE. 2016. IUCN SSC Guidelines for Assessing Species’ Vulnerability to Climate Change. Version 1.0. Occasional Paper of the IUCN Species Survival Commission No. 59. Cambridge, UK and Gland, Switzerland: IUCN Species Survival Commission. x+114pp.
Hart AG, Dawson M, Fourie R, MacTavish L, Goodenough AE. 2022. Comparing the effectiveness of camera trapping, driven transects and ad hoc records for surveying nocturnal mammals against a known species assemblage. Community Ecology 7:1-3.
Hey D. 1964. The control of vertebrate problem animals in the province of the Cape of Good Hope, Republic of South Africa. In: Proceedings of the Vertebrate Pest Conference. 2(2): 57-70.
Hofer, H. and Mills, M.G.L. 1998a. Worldwide distribution of Hyaenas. In: M.G.L. Mills and H. Hofer (eds), Hyaenas. Status Survey and Conservation Action Plan, pp. 39-63. IUCN/SSC Hyaena Specialist Group. IUCN, Gland, Switzerland and Cambridge, UK.
Kerley GI, Pressey RL, Cowling RM, Boshoff AF, Sims-Castley R. 2003. Options for the conservation of large and medium-sized mammals in the Cape Floristic Region hotspot, South Africa. Biological Conservation 112(1-2):169-190.
Kingdon, J. 1977. East African Mammals: An Atlas of Evolution in Africa. Volume IIIA (Carnivores). Academic Press, London, UK.
Koehler, C.E. and Richardson, P.R.K. 1990. Proteles cristatus. Mammalian Species 363: 1-6.
Koeppel KN, Van Schalkwyk OL, Thompson PN. 2022. Patterns of rabies cases in South Africa between 1993 and 2019, including the role of wildlife. Transboundary and Emerging Diseases. 69(2):836-848.
Koepfli KP, Jenks SM, Eizirik E, Zahirpour T, Van Valkenburgh B, Wayne RK. 2006. Molecular systematics of the Hyaenidae: relationships of a relictual lineage resolved by a molecular supermatrix. Molecular phylogenetics and evolution 38(3):603-20.
Kotze R, Bennett NC, Cameron EZ, de Vries JL, Marneweck DG, Pirk CW, Dalerum F. 2012. Temporal patterns of den use suggest polygamous mating patterns in an obligate monogamous mammal. Animal behaviour 84(6):1573-1578.
Kruuk H, Sands WA. 1972. The aardwolf (Proteles cristatus Sparrman) 1783 as predator of termites. African Journal of Ecology. 10(3):211-227.
Lloyd PH, Millar JCG. 1984. A questionnaire survey (1969-1974) of some of the larger mammals of the Cape Province. Bontebok 3: 1-49.
Loock D, SwanepoeL L. 2019. Assessing local patterns in carnivore occupancy and richness by camera trapping: a monitoring of diversity at Verloren Vallei Nature Reserve, Dullstroom, Mpumalanga. Wildlife & Resource Association (WRA), Bloemfontein.
Lynch, C.D. 1994. The mammals of Lesotho. Navorsinge van die Nasionale Museum Bloemfontein 10(4): 177-241.
Marneweck DG, Cameron EZ, Ganswindt A, Dalerum F. 2015. Behavioural and endocrine correlates to the aardwolf mating system. Mammalian Biology 80: 31–38.
Matsebula SN, Monadjem A, Roques KG, Garcelon DK. 2009. The diet of the aardwolf, Proteles cristatus at Malolotja Nature Reserve, western Swaziland. African Journal of Ecology 47: 448–451.
Monadjem A. 1998. Distributional patterns and conservation status of mammals of Swaziland, southern Africa. Koedoe 41(2): 45-59.
Nieman WA. 2018. Culture, Conflict, and Cuisine: A Quantitative Assessment of Terrestrial Vertebrate Off-Take at the HumanWildlife Interface. Msc thesis, University of Stellenbosch, Stellenbosch.
Nieman, W.A., Leslie, A.J. and Wilkinson, A., 2018. The Use of Animals and Animal-Derived Constituents in African Traditional Medicine and Other Cultural Applications: Townships in the Western Cape Province. Culture, Conflict and Cuisine: A Quantitative Assessment of Terrestrial Vertebrate Off-Take at the Human-Wildlife Interface, p.215.
Nieman WA, Schultz BC, Wilkinson A, Leslie AJ. 2021. Stakeholders’ perceptions of mammal occurrence and abundance on agricultural properties bordering the Boland Mountain Complex, South Africa. African Zoology 56(2): 104-116.
Nowak, R.M. 1991. Walker’s Mammals of the World. The Johns Hopkins University Press, Baltimore, USA and London, UK.
NWPG (North West Provincial Government) 2011. Bojanala Platinum District Mammal Report Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
NWPG (North West Provincial Government) 2012. Ngaka Modiri Molema District Mammal Report Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
NWPG (North West Provincial Government) 2013. Dr Ruth Segomotsi Mompata District Mammal Report Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
NWPG (North West Provincial Government) 2016. Report examining the status of small carnivores and the detriment of hunting them in the Molopo Bushveld of the Dr Ruth Segomotsi Mompati District of the North West. Department of Rural, Environment and Agricultural Development, North West Provincial Government, Mahikeng.
NWPG (North West Provincial Government) 2017. The status of free ranging carnivores in Bokone Bophirima and impacts of hunting them. Department of Rural, Environment and Agricultural Development, North West Provincial Government, Mahikeng.
NWPG (North West Provincial Government) 2024 The impact of roadkill on mammal populations in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Pietersen DW, Robertson MP. 2023. Dietary niche breadth and overlap of four sympatric southern African myrmecophagous mammal species, as inferred from the literature. African Zoology.58(2):29-38.
Power, R.J. 2014. The distribution and status of mammals in the North West Province. Department of Economic Development, Environment, Conservation & Tourism, North West Provincial Government, Mahikeng.
Richardson PRK. 1987. Food consumption and seasonal variation in the diet of the aardwolf Proteles cristatus in southern Africa. Zeitschrift für Säugetierkunde 52: 307–325.
Power RJ, Van Straaten A, Schaller R, Mooke M, Boshoff T, Nel HP. 2019. An inventory of mammals of the North West Province, South Africa. Annals of the Ditsong Museum of Natural History 8: 6-29.
Pretorius, M. 2019. Mesocarnivores in Protected Areas: ecological and anthropogenic determinants of habitat use in northern Kwa-Zulu Natal, South Africa. Msc thesis, University of Cape Town, Cape Town.
Pringle JA. 1977. The distribution of mammals in Natal. Part 2. Carnivora. Annals of the Natal Museum 23(1): 93-115.
Richardson, P.R.K. 1984. Socio-ecology of the aardwolf in relation to its conservation. Hyaena Specialist Group Newsletter 1: 32-36.
Richardson PRK. 1987a. Food Consumption and seasonal variation in the diet of the aardwolf Proteles cristatus in Southern Africa. Z. Saugetierk. 52: 307–325.
Richardson PR. 1987b. Aardwolf mating system: overt cuckoldry in an apparently monogamous mammal. South African Journal of Science 83(7):405-409.
Richardson PR, Coetzee M. 1988. Mate desertion in response to female promiscuity in the socially monogamous aardwolf Proteles cristatus. African Zoology 23(4):306-308.
Richardson PR. 1991. Territorial significance of scent marking during the non‐mating season in the aardwolf Proteles cristatus (Carnivora: Protelidae). Ethology 87(1‐2):9-27.
Richardson PR. 1993. The function of scent marking in territories: a resurrection of the intimidation hypothesis. Transactions of the Royal Society of South Africa 48(2):195-206.
Richardson PR, Levitan CD. 1994. Tolerance of aardwolves to defense secretions of Trinervitermes trinervoides. Journal of Mammalogy 75(1):84-91.
Richardson PR, Anderson MD. 2005. The physical and thermal characteristics of aardwolf dens. South African Journal of Wildlife Research 35(2):147-153.
Richmond-Coggan L. 2014. Comparative abundance and ranging behaviour of brown hyaena (Parahyaena brunnea) inside and outside protected areas in South Africa. Ph.D. Nottingham Trent University.
Schurch MP, McManus J, Goets S, Pardo LE, Gaynor D, Samuels I, Cupido C, Couldridge V, Smuts B. 2021. Wildlife-friendly livestock management promotes mammalian biodiversity recovery on a semi-arid Karoo farm in South Africa. Frontiers in Conservation Science 2:6. https://doi.org/10.3389/fcosc.2021.652415
Simelane TS, Kerley GI. 1998. Conservation implications of the use of vertebrates by Xhosa traditional healers in South Africa. South African Journal of Wildlife Research (4):121-126.
Skinner, J.D. and Chimimba, C.T. (eds). 2005. The Mammals of the Southern African Subregion. Cambridge University Press, United Kingdom, Cambridge.
Sliwa A. 1996. A functional analysis of scent marking and mating behaviour in the aardwolf, Proteles cristatus (Sparrman, 1783). University of Pretoria.
Sliwa A, Richardson PR. 1998. Responses of aardwolves, Proteles cristatus, Sparrman 1783, to translocated scent marks. Animal Behaviour. 56(1):137-146.
Smith K, Venter JA, Peel M, Keith M, Somers MJ. 2023. Temporal partitioning and the potential for avoidance behaviour within South African carnivore communities. Ecology and Evolution 13(8):e10380.
Spagnuolo OS, Lemerle MA, Holekamp KE, Wiesel I. 2022. The value of individual identification in studies of free-living hyenas and aardwolves. Mammalian Biology 102(4):1089-1112.
Stuart CT. 1981. Notes on the mammalian carnivores of the Cape Province. Bontebok 1: 1 – 58.
Van Jaarsveld AS. 1993. A comparative investigation of hyaenid and aardwolf life-histories, with notes on spotted hyaena mortality patterns. Transactions of the Royal Society of South Africa 48(2):219-232.
Van Jaarsveld AS, Richardson PR, Anderson MD. 1995. Post-natal growth and sustained lactational effort in the aardwolf: life-history implications. Functional Ecology.9:492-497.
Von Richter, W. 1972. Remarks on present distribution and abundance of some South African carnivores. South African Journal of Wildlife Research 2(1): 9-16.
Webster AB, Pretorius ME, Somers MJ. 2021. The determinants of mesocarnivore activity patterns in highveld grassland and riparian habitats. African Journal of Wildlife Research. 51(1):178-192.
Werdelin L, Kitchener AC, Abramov A, Veron G, San ED. 2021. The scientific name of the aardwolf is Proteles cristatus. African Journal of Wildlife Research 51(1):149-152.
Westbury MV, De Cahsan B, Dalerum F, Norén K, Hofreiter M. 2019. Aardwolf population diversity and phylogenetic positioning inferred using complete mitochondrial genomes. African Journal of Wildlife Research 49(1):27-33.
Westbury MV, Le Duc D, Duchêne DA, Krishnan A, Prost S, Rutschmann S, Grau JH, Dalén L, Weyrich A, Norén K, Werdelin L. 2021. Ecological specialization and evolutionary reticulation in extant hyaenidae. Molecular biology and evolution. 38(9):3884-3897.
Whiting MJ. Williams VL, Hibbitts TJ. 2016. A picture of health? Animal use and the Faraday traditional medicine market, South Africa. Journal of Ethnopharmacology. 179:265-73.
Wilkinson CE, Dheer A, Zett T, Torrents‐Ticó M, Yarnell RW, Bar Ziv E, Bhandari S, Jacobson A, Dloniak SM. 2024. Review of the global research on Hyaenidae and implications for conservation and management. Mammal Review 54(2):193-212.
Williams JB, Anderson MD, Richardson PR. 1997. Seasonal differences in field metabolism, water requirements, and foraging behavior of free‐living aardwolves. Ecology 78(8):2588-2602.
Woodgate Z, Drouilly M, Distiller G, O’Riain MJ. 2023. The effect of multi-use landscapes on mammal assemblages and its implication for conservation. Land 12(3):599. https://doi.org/10.3390/land12030599
Yalden, D.W., Largen, M.J., Kock, D. and Hillman, J.C. 1996. Catalogue of the mammals of Ethiopia and Eritrea. 7. Revised checklist, zoogeography and conservation. Tropical Zoology 9: 73-164.
Yarnell RW, MacTavish L. 2013. A novel record of aardwolf Proteles cristata feeding behaviour. Transactions of the Royal Society of South Africa 68: 81–82.
de Vries JL, Pirk CWW, Bateman PW, Dalerum F. 2011. Extension of the diet of an extreme foraging specialist, the aardwolf (Proteles cristata). African Zoology 46: 194–196.